This regulatory network was inferred from the input dataset. The miRNAs and mRNAs are
presented as round and rectangle nodes respectively. The numerical value popped up upon mouse over the gene node is the log2 transformed fold-change of the gene expression between the two groups. All of the nodes are clickable, and the detailed information of the miRNAs/mRNAs and related cancer pathway will be displayed in another window. The edges between nodes are supported by both interactions (predicted or experimentally verified) and correlations learnt from cancer dataset. The numerical value popped up upon mouse over the edge is the correlation beat value (effect size) between the two nodes. The experimental evidences of the edges reported in previous cancer studies are highlighted by red/orange color. All of these information can be accessed by the "mouse-over" action. This network shows a full map of the miRNA-mRNA regulation of the input gene list(s), and the hub miRNAs (with the high network degree/betweenness centrality) would be the potential cancer drivers or tumor suppressors. The full result table can be accessed in the "Regulations" tab.
"miRNACancerMAP" is also a network visualization tool for users to draw their regulatory network by personal customization. Users can set the complexity of the network by limiting the number of nodes or edges. And the color of the nodes can be defined by different categories of the mRNAs and miRNAs, such as Gene-Ontology, pathway, and expression status. Users can also select to use network degree or network betweenness centrality to define the node size. And edges can be black or colored by the correlation. Purple edge means negative correlation (mostly found between miRNA and mRNA), and blue edge means positive correlation (found in PPI or miRNA-miRNA sponge effect). We can also add the protein-protein interactions (PPI) into the network. This result will show the cluster of genes regulated by some specific miRNAs. Additionally, miRNA-miRNA edges can be added by the "miRNA sponge" button, presenting some clusters of miRNAs that have the interactions via sponge effect.
miRNA-gene regulations
| Num | microRNA | Gene | miRNA log2FC | miRNA pvalue | Gene log2FC | Gene pvalue | Interaction | Correlation beta | Correlation P-value | PMID | Reported in cancer studies |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | hsa-miR-30e-5p | ABL1 | 1.6 | 0 | -0.1 | 0.42908 | MirTarget; miRNATAP | -0.11 | 0.00016 | NA | |
| 2 | hsa-miR-140-3p | ANAPC1 | -1.11 | 0 | 0.43 | 0.00238 | MirTarget | -0.19 | 1.0E-5 | NA | |
| 3 | hsa-miR-26b-5p | ANAPC1 | 0.72 | 5.0E-5 | 0.43 | 0.00238 | miRNAWalker2 validate | -0.14 | 9.0E-5 | NA | |
| 4 | hsa-miR-30c-2-3p | ANAPC7 | -3.11 | 0 | 0.39 | 0.00036 | MirTarget | -0.13 | 0 | NA | |
| 5 | hsa-miR-664a-5p | ATR | -0.09 | 0.66227 | 0.55 | 0 | mirMAP | -0.1 | 9.0E-5 | NA | |
| 6 | hsa-miR-139-5p | BUB1 | -2.27 | 0 | 2.45 | 0 | miRanda | -0.44 | 0 | NA | |
| 7 | hsa-miR-199a-5p | BUB1 | 1.31 | 0 | 2.45 | 0 | miRanda | -0.18 | 0.0036 | NA | |
| 8 | hsa-miR-199b-5p | BUB1 | 2.14 | 0 | 2.45 | 0 | miRanda | -0.15 | 0.00543 | NA | |
| 9 | hsa-miR-486-5p | BUB1B | -4.39 | 0 | 3.44 | 0 | miRanda | -0.28 | 0 | NA | |
| 10 | hsa-miR-139-5p | BUB3 | -2.27 | 0 | 0.63 | 0 | miRanda | -0.14 | 0 | NA | |
| 11 | hsa-miR-107 | CCNA1 | 0.66 | 0 | -1.86 | 0.00094 | miRanda | -0.63 | 0.00151 | NA | |
| 12 | hsa-miR-30d-5p | CCNA1 | -0.92 | 4.0E-5 | -1.86 | 0.00094 | MirTarget | -0.3 | 0.00946 | NA | |
| 13 | hsa-let-7a-3p | CCNA2 | 0.17 | 0.43183 | 2.69 | 0 | MirTarget | -0.23 | 0.00207 | NA | |
| 14 | hsa-let-7b-3p | CCNA2 | -1.82 | 0 | 2.69 | 0 | MirTarget | -0.53 | 0 | NA | |
| 15 | hsa-let-7b-5p | CCNA2 | -1.62 | 0 | 2.69 | 0 | miRNAWalker2 validate; miRTarBase | -0.59 | 0 | NA | |
| 16 | hsa-miR-200b-3p | CCNA2 | 1.55 | 0 | 2.69 | 0 | TargetScan | -0.2 | 0.00018 | NA | |
| 17 | hsa-miR-218-5p | CCNA2 | -0.28 | 0.25955 | 2.69 | 0 | MirTarget | -0.39 | 0 | NA | |
| 18 | hsa-miR-27b-3p | CCNA2 | 0.24 | 0.12264 | 2.69 | 0 | miRNATAP | -0.36 | 0.00016 | NA | |
| 19 | hsa-miR-29a-3p | CCNA2 | 0.1 | 0.5732 | 2.69 | 0 | MirTarget | -0.46 | 0 | NA | |
| 20 | hsa-miR-29c-3p | CCNA2 | 1.32 | 0 | 2.69 | 0 | MirTarget | -0.3 | 0 | NA | |
| 21 | hsa-miR-320a | CCNA2 | -0.96 | 0 | 2.69 | 0 | miRanda | -0.33 | 7.0E-5 | NA | |
| 22 | hsa-miR-34c-5p | CCNA2 | -1 | 0.07244 | 2.69 | 0 | miRanda | -0.14 | 0 | NA | |
| 23 | hsa-miR-455-5p | CCNA2 | 1.37 | 0 | 2.69 | 0 | miRanda | -0.25 | 9.0E-5 | NA | |
| 24 | hsa-miR-486-5p | CCNA2 | -4.39 | 0 | 2.69 | 0 | miRanda | -0.22 | 0 | NA | |
| 25 | hsa-miR-508-3p | CCNA2 | 0.14 | 0.79184 | 2.69 | 0 | MirTarget; PITA; miRNATAP | -0.14 | 0 | NA | |
| 26 | hsa-let-7b-5p | CCNB1 | -1.62 | 0 | 2.59 | 0 | miRNAWalker2 validate | -0.6 | 0 | NA | |
| 27 | hsa-miR-139-5p | CCNB1 | -2.27 | 0 | 2.59 | 0 | miRanda | -0.4 | 0 | NA | |
| 28 | hsa-let-7a-5p | CCNB2 | -1.37 | 0 | 3.05 | 0 | miRNAWalker2 validate | -0.57 | 0 | NA | |
| 29 | hsa-let-7b-5p | CCNB2 | -1.62 | 0 | 3.05 | 0 | miRNAWalker2 validate | -0.61 | 0 | NA | |
| 30 | hsa-let-7c-5p | CCNB2 | -2.14 | 0 | 3.05 | 0 | miRNAWalker2 validate | -0.62 | 0 | NA | |
| 31 | hsa-miR-23b-3p | CCNB2 | -0.07 | 0.62059 | 3.05 | 0 | miRNAWalker2 validate | -0.6 | 0 | NA | |
| 32 | hsa-miR-106a-5p | CCND1 | 1.39 | 6.0E-5 | -0.3 | 0.2554 | MirTarget; miRNATAP | -0.25 | 0 | NA | |
| 33 | hsa-miR-106b-5p | CCND1 | 1.47 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase; MirTarget; miRNATAP | -0.26 | 1.0E-5 | NA | |
| 34 | hsa-miR-142-3p | CCND1 | 3.98 | 0 | -0.3 | 0.2554 | miRanda | -0.12 | 0.00053 | 23619912 | Transfection of miR-142-3p mimics in colon cancer cells downregulated cyclin D1 expression induced G1 phase cell cycle arrest and elevated the sensitivity of the cells to 5-fluorouracil |
| 35 | hsa-miR-15a-5p | CCND1 | 1.63 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase; MirTarget; miRNATAP | -0.2 | 0.00193 | 22922827 | CCND1 has been found to be a target of miR-15a and miR-16-1 through analysis of complementary sequences between microRNAs and CCND1 mRNA; Moreover the transcription of CCND1 is suppressed by miR-15a and miR-16-1 via direct binding to the CCND1 3'-untranslated region 3'-UTR |
| 36 | hsa-miR-15b-5p | CCND1 | -1.26 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase; MirTarget; miRNATAP | -0.3 | 0 | NA | |
| 37 | hsa-miR-16-1-3p | CCND1 | 1.5 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase | -0.25 | 3.0E-5 | 22922827; 18483394 | CCND1 has been found to be a target of miR-15a and miR-16-1 through analysis of complementary sequences between microRNAs and CCND1 mRNA; Moreover the transcription of CCND1 is suppressed by miR-15a and miR-16-1 via direct binding to the CCND1 3'-untranslated region 3'-UTR;Truncation in CCND1 mRNA alters miR 16 1 regulation in mantle cell lymphoma; Furthermore we demonstrated that this truncation alters miR-16-1 binding sites and through the use of reporter constructs we were able to show that miR-16-1 regulates CCND1 mRNA expression; This study introduces the role of miR-16-1 in the regulation of CCND1 in MCL |
| 38 | hsa-miR-16-5p | CCND1 | 0.75 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase; MirTarget; miRNATAP | -0.37 | 0 | 23991964; 22922827; 18483394 | At the molecular level our results further revealed that cyclin D1 expression was negatively regulated by miR-16;CCND1 has been found to be a target of miR-15a and miR-16-1 through analysis of complementary sequences between microRNAs and CCND1 mRNA; Moreover the transcription of CCND1 is suppressed by miR-15a and miR-16-1 via direct binding to the CCND1 3'-untranslated region 3'-UTR;Truncation in CCND1 mRNA alters miR 16 1 regulation in mantle cell lymphoma; Furthermore we demonstrated that this truncation alters miR-16-1 binding sites and through the use of reporter constructs we were able to show that miR-16-1 regulates CCND1 mRNA expression; This study introduces the role of miR-16-1 in the regulation of CCND1 in MCL |
| 39 | hsa-miR-17-5p | CCND1 | 2.07 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; MirTarget; TargetScan; miRNATAP | -0.31 | 0 | 26431674 | Bioinformatics Prediction and In Vitro Analysis Revealed That miR 17 Targets Cyclin D1 mRNA in Triple Negative Breast Cancer Cells; In this study using bioinformatic analyses miR-17 was selected as it targets the 3'UTR of CCND1 gene with the highest score; After lentiviral transduction of miR-17 to the target cells gene expression analysis showed decreased expression of CCND1 gene |
| 40 | hsa-miR-186-5p | CCND1 | 0.85 | 0 | -0.3 | 0.2554 | mirMAP | -0.38 | 1.0E-5 | NA | |
| 41 | hsa-miR-193a-3p | CCND1 | 0.55 | 0.0319 | -0.3 | 0.2554 | MirTarget; PITA; miRanda | -0.19 | 0.00016 | NA | |
| 42 | hsa-miR-193b-3p | CCND1 | 1.1 | 0.00082 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.15 | 4.0E-5 | 27071318; 20655737; 20304954; 21893020; 26129688 | MicroRNA 193b inhibits the proliferation migration and invasion of gastric cancer cells via targeting cyclin D1; Further mechanism study indicated that CCND1 was a direct target of miR-193b in GC;CCND1 and ETS1 were revealed to be regulated by miR-193b directly;MicroRNA 193b represses cell proliferation and regulates cyclin D1 in melanoma; Overexpression of miR-193b in Malme-3M cells down-regulated CCND1 mRNA and protein by > or = 50%; A luciferase reporter assay confirmed that miR-193b directly regulates CCND1 by binding to the 3'untranslated region of CCND1 mRNA; These studies indicate that miR-193b represses cell proliferation and regulates CCND1 expression and suggest that dysregulation of miR-193b may play an important role in melanoma development;In a previous study we reported that miR-193b represses cell proliferation and regulates cyclin D1 in melanoma cells suggesting that miR-193b could act as a tumor suppressor;Epigenetically altered miR 193b targets cyclin D1 in prostate cancer; It has been suggested that miR-193b targets cyclin D1 in several malignancies; Here our aim was to determine if miR-193b targets cyclin D1 in prostate cancer; Furthermore the PC cell lines 22Rv1 and VCaP which express low levels of miR-193b and high levels of CCND1 showed significant growth retardation when treated with a CDK4/6 inhibitor; In contrast the inhibitor had no effect on the growth of PC-3 and DU145 cells with high miR-193b and low CCND1 expression; Taken together our data demonstrate that miR-193b targets cyclin D1 in prostate cancer |
| 43 | hsa-miR-19a-3p | CCND1 | 2.12 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.22 | 0 | 25985117 | Moreover miR-19a might play inhibitory roles in HCC malignancy via regulating Cyclin D1 expression |
| 44 | hsa-miR-19b-1-5p | CCND1 | 1.71 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase | -0.3 | 0 | NA | |
| 45 | hsa-miR-19b-3p | CCND1 | 2.11 | 0 | -0.3 | 0.2554 | miRNATAP | -0.18 | 0.0002 | NA | |
| 46 | hsa-miR-20a-5p | CCND1 | 2.65 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase; MirTarget; miRNATAP | -0.24 | 0 | NA | |
| 47 | hsa-miR-20b-5p | CCND1 | 1.36 | 0.00261 | -0.3 | 0.2554 | MirTarget; miRNATAP | -0.18 | 0 | NA | |
| 48 | hsa-miR-365a-3p | CCND1 | 0.01 | 0.9536 | -0.3 | 0.2554 | miRNAWalker2 validate; miRTarBase | -0.19 | 0.00023 | NA | |
| 49 | hsa-miR-374a-5p | CCND1 | -0.2 | 0.29808 | -0.3 | 0.2554 | MirTarget | -0.31 | 1.0E-5 | 27191497 | microRNA 374a suppresses colon cancer progression by directly reducing CCND1 to inactivate the PI3K/AKT pathway; Furthermore luciferase reporter assays confirmed that miR-374a could directly reduce CCND1; We examined miR-374a levels by in situ hybridization and its correlation with CCND1 expression in CRC tumor tissues; High miR-374a expression with low level of CCND1 was protective factor in CRC; Together these findings indicate that miR-374a inactivates the PI3K/AKT axis by inhibiting CCND1 suppressing of colon cancer progression |
| 50 | hsa-miR-374b-5p | CCND1 | 0.47 | 0.01092 | -0.3 | 0.2554 | miRNAWalker2 validate; MirTarget | -0.26 | 0.00012 | NA | |
| 51 | hsa-miR-425-5p | CCND1 | 1.22 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate | -0.25 | 1.0E-5 | NA | |
| 52 | hsa-miR-589-3p | CCND1 | 1.34 | 2.0E-5 | -0.3 | 0.2554 | MirTarget | -0.13 | 0.00079 | NA | |
| 53 | hsa-miR-590-3p | CCND1 | 0.84 | 0.00129 | -0.3 | 0.2554 | mirMAP | -0.18 | 0.00086 | NA | |
| 54 | hsa-miR-769-3p | CCND1 | 0.45 | 0.07482 | -0.3 | 0.2554 | mirMAP | -0.14 | 0.00277 | NA | |
| 55 | hsa-miR-92a-3p | CCND1 | -0.14 | 0.49341 | -0.3 | 0.2554 | miRNAWalker2 validate | -0.34 | 0 | NA | |
| 56 | hsa-miR-93-5p | CCND1 | 1.51 | 0 | -0.3 | 0.2554 | miRNAWalker2 validate; MirTarget; miRNATAP | -0.32 | 0 | NA | |
| 57 | hsa-miR-942-5p | CCND1 | -0.04 | 0.87063 | -0.3 | 0.2554 | MirTarget | -0.18 | 0.00098 | NA | |
| 58 | hsa-miR-106a-5p | CCND2 | 1.39 | 6.0E-5 | -1.64 | 0 | miRNATAP | -0.15 | 2.0E-5 | NA | |
| 59 | hsa-miR-106b-5p | CCND2 | 1.47 | 0 | -1.64 | 0 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.38 | 0 | NA | |
| 60 | hsa-miR-130b-5p | CCND2 | 1.54 | 0 | -1.64 | 0 | mirMAP | -0.27 | 0 | NA | |
| 61 | hsa-miR-141-3p | CCND2 | 3.37 | 0 | -1.64 | 0 | MirTarget; TargetScan | -0.24 | 0 | NA | |
| 62 | hsa-miR-151a-3p | CCND2 | 0.67 | 0.00028 | -1.64 | 0 | mirMAP | -0.28 | 0 | NA | |
| 63 | hsa-miR-15a-5p | CCND2 | 1.63 | 0 | -1.64 | 0 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.16 | 0.00712 | NA | |
| 64 | hsa-miR-17-5p | CCND2 | 2.07 | 0 | -1.64 | 0 | miRNAWalker2 validate; miRTarBase; TargetScan; miRNATAP | -0.37 | 0 | NA | |
| 65 | hsa-miR-182-5p | CCND2 | 3.22 | 0 | -1.64 | 0 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.37 | 0 | NA | |
| 66 | hsa-miR-183-5p | CCND2 | 2.39 | 0 | -1.64 | 0 | miRNATAP | -0.41 | 0 | NA | |
| 67 | hsa-miR-185-5p | CCND2 | 1.14 | 0 | -1.64 | 0 | MirTarget; miRNATAP | -0.24 | 0.00048 | NA | |
| 68 | hsa-miR-186-5p | CCND2 | 0.85 | 0 | -1.64 | 0 | mirMAP; miRNATAP | -0.36 | 1.0E-5 | NA | |
| 69 | hsa-miR-191-5p | CCND2 | 0.34 | 0.06681 | -1.64 | 0 | MirTarget | -0.2 | 0.00075 | NA | |
| 70 | hsa-miR-19a-3p | CCND2 | 2.12 | 0 | -1.64 | 0 | MirTarget; miRNATAP | -0.17 | 1.0E-5 | NA | |
| 71 | hsa-miR-19b-3p | CCND2 | 2.11 | 0 | -1.64 | 0 | miRNAWalker2 validate; MirTarget; miRNATAP | -0.19 | 1.0E-5 | NA | |
| 72 | hsa-miR-200a-3p | CCND2 | 3.15 | 0 | -1.64 | 0 | MirTarget | -0.17 | 0 | NA | |
| 73 | hsa-miR-20a-5p | CCND2 | 2.65 | 0 | -1.64 | 0 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.28 | 0 | NA | |
| 74 | hsa-miR-21-3p | CCND2 | 2.54 | 0 | -1.64 | 0 | mirMAP | -0.21 | 0 | NA | |
| 75 | hsa-miR-2355-3p | CCND2 | 1.11 | 1.0E-5 | -1.64 | 0 | miRNATAP | -0.22 | 0 | NA | |
| 76 | hsa-miR-28-5p | CCND2 | 1.2 | 0 | -1.64 | 0 | miRanda | -0.29 | 0.00018 | NA | |
| 77 | hsa-miR-301a-3p | CCND2 | 2.7 | 0 | -1.64 | 0 | miRNAWalker2 validate | -0.27 | 0 | NA | |
| 78 | hsa-miR-320b | CCND2 | 0.23 | 0.37882 | -1.64 | 0 | mirMAP; miRNATAP | -0.12 | 0.00373 | NA | |
| 79 | hsa-miR-324-3p | CCND2 | -0.08 | 0.68923 | -1.64 | 0 | miRNAWalker2 validate | -0.28 | 0 | NA | |
| 80 | hsa-miR-331-5p | CCND2 | 0.58 | 0.00131 | -1.64 | 0 | miRNATAP | -0.28 | 1.0E-5 | NA | |
| 81 | hsa-miR-429 | CCND2 | 2.38 | 0 | -1.64 | 0 | miRNATAP | -0.21 | 0 | NA | |
| 82 | hsa-miR-450b-5p | CCND2 | 1.69 | 0 | -1.64 | 0 | MirTarget; PITA; miRNATAP | -0.15 | 9.0E-5 | NA | |
| 83 | hsa-miR-501-5p | CCND2 | 0.41 | 0.10435 | -1.64 | 0 | PITA; mirMAP; miRNATAP | -0.14 | 0.00086 | NA | |
| 84 | hsa-miR-503-5p | CCND2 | 1.97 | 0 | -1.64 | 0 | MirTarget | -0.13 | 5.0E-5 | 25860935 | We then identified two targets of miR-503 CCND2 and CCND3 |
| 85 | hsa-miR-550a-5p | CCND2 | 0.6 | 0.03148 | -1.64 | 0 | MirTarget | -0.22 | 0 | NA | |
| 86 | hsa-miR-589-3p | CCND2 | 1.34 | 2.0E-5 | -1.64 | 0 | mirMAP | -0.19 | 0 | NA | |
| 87 | hsa-miR-590-3p | CCND2 | 0.84 | 0.00129 | -1.64 | 0 | miRanda; mirMAP | -0.13 | 0.00821 | NA | |
| 88 | hsa-miR-590-5p | CCND2 | 2.07 | 0 | -1.64 | 0 | mirMAP | -0.26 | 0 | NA | |
| 89 | hsa-miR-660-5p | CCND2 | 2.05 | 0 | -1.64 | 0 | mirMAP | -0.15 | 0.00131 | NA | |
| 90 | hsa-miR-7-1-3p | CCND2 | 2.61 | 0 | -1.64 | 0 | mirMAP | -0.26 | 0 | NA | |
| 91 | hsa-miR-877-5p | CCND2 | -0.37 | 0.20671 | -1.64 | 0 | miRNAWalker2 validate | -0.15 | 6.0E-5 | NA | |
| 92 | hsa-miR-93-5p | CCND2 | 1.51 | 0 | -1.64 | 0 | miRNATAP | -0.41 | 0 | NA | |
| 93 | hsa-miR-96-5p | CCND2 | 3.04 | 0 | -1.64 | 0 | TargetScan; miRNATAP | -0.36 | 0 | NA | |
| 94 | hsa-miR-320b | CCND3 | 0.23 | 0.37882 | -0.86 | 1.0E-5 | miRanda | -0.11 | 0.00235 | NA | |
| 95 | hsa-miR-421 | CCND3 | 0.17 | 0.53528 | -0.86 | 1.0E-5 | PITA; miRanda | -0.12 | 0.00073 | NA | |
| 96 | hsa-miR-96-5p | CCND3 | 3.04 | 0 | -0.86 | 1.0E-5 | TargetScan | -0.12 | 0.00015 | NA | |
| 97 | hsa-miR-125b-5p | CCNE1 | -0.55 | 0.01072 | 3 | 0 | miRNAWalker2 validate | -0.21 | 0.00408 | NA | |
| 98 | hsa-miR-195-5p | CCNE1 | -1.02 | 5.0E-5 | 3 | 0 | miRNAWalker2 validate; MirTarget; miRNATAP | -0.52 | 0 | 24402230 | Furthermore through qPCR and western blot assays we showed that overexpression of miR-195-5p reduced CCNE1 mRNA and protein levels respectively |
| 99 | hsa-miR-26a-5p | CCNE1 | -0.13 | 0.44003 | 3 | 0 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.7 | 0 | 22094936 | Cell cycle regulation and CCNE1 and CDC2 were the only significant overlapping pathway and genes differentially expressed between tumors with high and low levels of miR-26a and EZH2 respectively; Low mRNA levels of EZH2 CCNE1 and CDC2 and high levels of miR-26a are associated with favorable outcome on tamoxifen |
| 100 | hsa-miR-497-5p | CCNE1 | -0.05 | 0.78621 | 3 | 0 | MirTarget; miRNATAP | -0.5 | 0 | 25909221; 24112607; 24909281 | The effect of simultaneous overexpression of miR-497 and miR-34a on the inhibition of cell proliferation colony formation and tumor growth and the downregulation of cyclin E1 was stronger than the effect of each miRNA alone; The synergistic actions of miR-497 and miR-34a partly correlated with cyclin E1 levels; These results indicate cyclin E1 is downregulated by both miR-497 and miR-34a which synergistically retard the growth of human lung cancer cells;Western blot assays confirmed that overexpression of miR-497 reduced cyclin E1 protein levels; Inhibited cellular growth suppressed cellular migration and invasion and G1 cell cycle arrest were observed upon overexpression of miR-497 in cells possibly by targeting cyclin E1;miR 497 suppresses proliferation of human cervical carcinoma HeLa cells by targeting cyclin E1; Furthermore the target effect of miR-497 on the CCNE1 was identified by dual-luciferase reporter assay system qRT-PCR and Western blotting; Over-expressed miR-497 in HeLa cells could suppress cell proliferation by targeting CCNE1 |
| 101 | hsa-let-7a-3p | CCNE2 | 0.17 | 0.43183 | 2.15 | 0 | mirMAP | -0.22 | 0.00307 | NA | |
| 102 | hsa-let-7b-3p | CCNE2 | -1.82 | 0 | 2.15 | 0 | mirMAP | -0.49 | 0 | NA | |
| 103 | hsa-miR-126-3p | CCNE2 | 0.4 | 0.11564 | 2.15 | 0 | miRNAWalker2 validate | -0.15 | 0.00656 | NA | |
| 104 | hsa-miR-26a-5p | CCNE2 | -0.13 | 0.44003 | 2.15 | 0 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.51 | 0 | 24116110; 21901171 | The loss of miR 26a mediated post transcriptional regulation of cyclin E2 in pancreatic cancer cell proliferation and decreased patient survival; The in vitro and in vivo assays showed that overexpression of miR-26a resulted in cell cycle arrest inhibited cell proliferation and decreased tumor growth which was associated with cyclin E2 downregulation;We also show that enforced expression of miR-26a in AML cells is able to inhibit cell cycle progression by downregulating cyclin E2 expression |
| 105 | hsa-miR-26b-5p | CCNE2 | 0.72 | 5.0E-5 | 2.15 | 0 | miRNATAP | -0.22 | 0.00477 | NA | |
| 106 | hsa-miR-3065-5p | CCNE2 | 0.65 | 0.09995 | 2.15 | 0 | mirMAP | -0.2 | 0 | NA | |
| 107 | hsa-miR-30a-5p | CCNE2 | -0.92 | 0.00076 | 2.15 | 0 | miRNATAP | -0.49 | 0 | NA | |
| 108 | hsa-miR-30b-5p | CCNE2 | 0.36 | 0.13803 | 2.15 | 0 | miRNAWalker2 validate; miRTarBase | -0.2 | 0.0005 | 22384020 | A luciferase-based reporter assay demonstrated that miR-30b post-transcriptionally reduced 27% p = 0.005 of the gene expression by interacting with two binding sites in the 3'-UTR of CCNE2; The upregulation of miR-30b by trastuzumab may play a biological role in trastuzumab-induced cell growth inhibition by targeting CCNE2 |
| 109 | hsa-miR-30c-5p | CCNE2 | -0.33 | 0.1236 | 2.15 | 0 | miRNATAP | -0.44 | 0 | NA | |
| 110 | hsa-miR-30d-5p | CCNE2 | -0.92 | 4.0E-5 | 2.15 | 0 | miRNATAP | -0.35 | 0 | 25843294 | MicroRNA 30d 5p inhibits tumour cell proliferation and motility by directly targeting CCNE2 in non small cell lung cancer; In addition the re-introduction of CCNE2 expression antagonised the inhibitory effects of miR-30d-5p on the capacity of NSCLC cells for proliferation and motility |
| 111 | hsa-miR-34a-5p | CCNE2 | 1.41 | 0 | 2.15 | 0 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.2 | 0.00337 | NA | |
| 112 | hsa-miR-34c-5p | CCNE2 | -1 | 0.07244 | 2.15 | 0 | miRNAWalker2 validate; miRTarBase; PITA; miRanda; miRNATAP | -0.11 | 4.0E-5 | NA | |
| 113 | hsa-miR-514a-3p | CCNE2 | -0.62 | 0.31374 | 2.15 | 0 | miRNATAP | -0.17 | 0 | NA | |
| 114 | hsa-miR-1301-3p | CCNH | 0.85 | 0.00058 | -0.17 | 0.11937 | miRNAWalker2 validate | -0.13 | 0 | NA | |
| 115 | hsa-miR-141-3p | CDC14A | 3.37 | 0 | -1.19 | 0 | TargetScan; miRNATAP | -0.19 | 0 | NA | |
| 116 | hsa-miR-324-5p | CDC14A | 1.07 | 5.0E-5 | -1.19 | 0 | miRanda | -0.12 | 0.00048 | NA | |
| 117 | hsa-miR-576-5p | CDC14A | 1.03 | 0 | -1.19 | 0 | mirMAP | -0.13 | 0.00347 | NA | |
| 118 | hsa-miR-590-5p | CDC14B | 2.07 | 0 | -0.34 | 0.01664 | miRanda | -0.1 | 0.00044 | NA | |
| 119 | hsa-miR-23b-3p | CDC20 | -0.07 | 0.62059 | 3.63 | 0 | miRNAWalker2 validate | -0.66 | 0 | NA | |
| 120 | hsa-miR-30a-5p | CDC20 | -0.92 | 0.00076 | 3.63 | 0 | miRNAWalker2 validate | -0.66 | 0 | NA | |
| 121 | hsa-let-7a-5p | CDC25A | -1.37 | 0 | 2.44 | 0 | MirTarget; TargetScan; miRNATAP | -0.53 | 0 | NA | |
| 122 | hsa-let-7b-5p | CDC25A | -1.62 | 0 | 2.44 | 0 | miRNAWalker2 validate; miRTarBase; MirTarget; miRNATAP | -0.58 | 0 | NA | |
| 123 | hsa-let-7c-5p | CDC25A | -2.14 | 0 | 2.44 | 0 | MirTarget | -0.55 | 0 | 25909324 | MicroRNA let 7c Inhibits Cell Proliferation and Induces Cell Cycle Arrest by Targeting CDC25A in Human Hepatocellular Carcinoma; The aim of the present study was to determine whether the cell cycle regulator CDC25A is involved in the antitumor effect of let-7c in HCC; The luciferase reporter assay showed that CDC25A was a direct target of let-7c and that let-7c inhibited the expression of CDC25A protein by directly targeting its 3' UTR; In conclusion this study indicates that let-7c suppresses HCC progression possibly by directly targeting the cell cycle regulator CDC25A and indirectly affecting its downstream target molecules |
| 124 | hsa-let-7f-5p | CDC25A | -0.05 | 0.83408 | 2.44 | 0 | MirTarget; miRNATAP | -0.17 | 0.00273 | NA | |
| 125 | hsa-miR-195-5p | CDC25A | -1.02 | 5.0E-5 | 2.44 | 0 | MirTarget; miRNATAP | -0.47 | 0 | NA | |
| 126 | hsa-miR-30b-3p | CDC25A | -1.21 | 0 | 2.44 | 0 | MirTarget | -0.31 | 0 | NA | |
| 127 | hsa-miR-34c-5p | CDC25A | -1 | 0.07244 | 2.44 | 0 | miRNATAP | -0.15 | 0 | 21321636 | Ectopic expression of miR-449b and miR-34c resulted in lowered adhesion activities by 28%-34% and in cell cycle arrests with increased cell number of 15.62% and 15.71% in G1 and with decreased cell number of 15.96% and 16.56% in S Cell cycle related proteins CDK6 and CDC25A were down-regulated; The decreases of CDK6 and CDC25A by miR-449b were 39% and 22% respecyively; 49% and 32% by miR-34c respectively |
| 128 | hsa-miR-497-5p | CDC25A | -0.05 | 0.78621 | 2.44 | 0 | MirTarget; miRNATAP | -0.56 | 0 | NA | |
| 129 | hsa-let-7a-5p | CDC25B | -1.37 | 0 | 0.43 | 0.03249 | miRNAWalker2 validate | -0.28 | 0 | NA | |
| 130 | hsa-miR-200b-3p | CDC27 | 1.55 | 0 | 0.08 | 0.43114 | TargetScan | -0.11 | 0 | NA | |
| 131 | hsa-miR-362-5p | CDC27 | 0.66 | 0.02433 | 0.08 | 0.43114 | miRNATAP | -0.11 | 0 | NA | |
| 132 | hsa-miR-497-5p | CDC27 | -0.05 | 0.78621 | 0.08 | 0.43114 | miRNATAP | -0.2 | 0 | NA | |
| 133 | hsa-miR-26a-5p | CDC6 | -0.13 | 0.44003 | 3.07 | 0 | miRNAWalker2 validate | -0.73 | 0 | 25100863; 27158389 | Here it is demonstrated that miR26a and miR26b inhibit replication licensing and the proliferation migration and invasion of lung cancer cells by targeting CDC6; The current study suggests that miR26a miR26b and CDC6 and factors regulating their expression represent potential cancer diagnostic and prognostic markers as well as anticancer targets;miR 26a inhibits the proliferation of ovarian cancer cells via regulating CDC6 expression; Bioinformatics analysis revealed Cdc6 was a target gene of miR-26a; dual-luciferase assay and validation assay showed miR-26a could act on the 3'UTR of Cdc6 to regulate Cdc6 expression; These findings suggest that miR-26a may act on the 3'UTR of Cdc6 to regulate Cdc6 expression which then inhibit the proliferation of ovarian cancer cells and induce their apoptosis |
| 134 | hsa-miR-361-3p | CDC6 | 0.1 | 0.54913 | 3.07 | 0 | miRNAWalker2 validate | -0.41 | 2.0E-5 | NA | |
| 135 | hsa-let-7a-5p | CDC7 | -1.37 | 0 | 1.27 | 0 | miRNAWalker2 validate | -0.32 | 0 | NA | |
| 136 | hsa-miR-3065-3p | CDC7 | 0.38 | 0.30796 | 1.27 | 0 | MirTarget | -0.18 | 0 | NA | |
| 137 | hsa-miR-338-5p | CDC7 | -1.83 | 0 | 1.27 | 0 | mirMAP | -0.19 | 0 | NA | |
| 138 | hsa-miR-455-5p | CDC7 | 1.37 | 0 | 1.27 | 0 | miRanda | -0.17 | 0.00042 | NA | |
| 139 | hsa-miR-23b-3p | CDK2 | -0.07 | 0.62059 | 0.34 | 0.01969 | miRNAWalker2 validate | -0.32 | 0 | NA | |
| 140 | hsa-miR-26b-5p | CDK2 | 0.72 | 5.0E-5 | 0.34 | 0.01969 | miRNAWalker2 validate | -0.13 | 0.00058 | NA | |
| 141 | hsa-miR-145-5p | CDK4 | -1.35 | 0 | 0.74 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.15 | 0 | 21092188 | Furthermore we found that CDK4 was regulated by miR-145 in cell cycle control |
| 142 | hsa-miR-195-5p | CDK4 | -1.02 | 5.0E-5 | 0.74 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.11 | 0.00037 | NA | |
| 143 | hsa-miR-320a | CDK4 | -0.96 | 0 | 0.74 | 1.0E-5 | miRNAWalker2 validate | -0.2 | 1.0E-5 | NA | |
| 144 | hsa-miR-141-3p | CDK6 | 3.37 | 0 | -0.63 | 0.01747 | TargetScan; miRNATAP | -0.16 | 6.0E-5 | NA | |
| 145 | hsa-miR-148a-3p | CDK6 | 2.31 | 0 | -0.63 | 0.01747 | mirMAP | -0.14 | 0.00378 | NA | |
| 146 | hsa-miR-15a-5p | CDK6 | 1.63 | 0 | -0.63 | 0.01747 | miRNATAP | -0.21 | 0.00126 | NA | |
| 147 | hsa-miR-16-1-3p | CDK6 | 1.5 | 0 | -0.63 | 0.01747 | mirMAP | -0.19 | 0.0025 | NA | |
| 148 | hsa-miR-182-5p | CDK6 | 3.22 | 0 | -0.63 | 0.01747 | mirMAP | -0.2 | 2.0E-5 | NA | |
| 149 | hsa-miR-200a-3p | CDK6 | 3.15 | 0 | -0.63 | 0.01747 | miRNATAP | -0.17 | 0 | 24009066 | microRNA 200a is an independent prognostic factor of hepatocellular carcinoma and induces cell cycle arrest by targeting CDK6 |
| 150 | hsa-miR-200b-3p | CDK6 | 1.55 | 0 | -0.63 | 0.01747 | mirMAP | -0.17 | 0.00022 | NA |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | CELL CYCLE | 74 | 1316 | 5.307e-76 | 2.469e-72 |
| 2 | CELL CYCLE PROCESS | 69 | 1081 | 1.548e-72 | 3.602e-69 |
| 3 | MITOTIC CELL CYCLE | 62 | 766 | 1.92e-69 | 2.978e-66 |
| 4 | REGULATION OF CELL CYCLE | 63 | 949 | 2.544e-65 | 2.959e-62 |
| 5 | CELL CYCLE PHASE TRANSITION | 38 | 255 | 7.328e-50 | 6.82e-47 |
| 6 | REGULATION OF MITOTIC CELL CYCLE | 43 | 468 | 1.474e-47 | 1.143e-44 |
| 7 | REGULATION OF CELL CYCLE PROCESS | 42 | 558 | 1.229e-42 | 8.172e-40 |
| 8 | REGULATION OF CELL CYCLE PHASE TRANSITION | 36 | 321 | 2.101e-42 | 1.141e-39 |
| 9 | NEGATIVE REGULATION OF CELL CYCLE | 39 | 433 | 2.207e-42 | 1.141e-39 |
| 10 | CELL DIVISION | 38 | 460 | 9.372e-40 | 4.361e-37 |
| 11 | CELL CYCLE CHECKPOINT | 30 | 194 | 2.075e-39 | 8.776e-37 |
| 12 | NEGATIVE REGULATION OF CELL CYCLE PROCESS | 30 | 214 | 4.708e-38 | 1.826e-35 |
| 13 | CELL CYCLE G1 S PHASE TRANSITION | 25 | 111 | 3.745e-37 | 1.245e-34 |
| 14 | G1 S TRANSITION OF MITOTIC CELL CYCLE | 25 | 111 | 3.745e-37 | 1.245e-34 |
| 15 | NEGATIVE REGULATION OF MITOTIC CELL CYCLE | 28 | 199 | 1.713e-35 | 5.312e-33 |
| 16 | MITOTIC CELL CYCLE CHECKPOINT | 24 | 139 | 1.267e-32 | 3.684e-30 |
| 17 | REGULATION OF TRANSFERASE ACTIVITY | 40 | 946 | 1.671e-30 | 4.573e-28 |
| 18 | NEGATIVE REGULATION OF CELL CYCLE PHASE TRANSITION | 23 | 146 | 2.801e-30 | 7.24e-28 |
| 19 | MITOTIC NUCLEAR DIVISION | 29 | 361 | 1.592e-29 | 3.898e-27 |
| 20 | REGULATION OF PROTEIN MODIFICATION PROCESS | 48 | 1710 | 2.083e-29 | 4.847e-27 |
| 21 | POSITIVE REGULATION OF CELL CYCLE | 28 | 332 | 4.504e-29 | 9.98e-27 |
| 22 | DNA INTEGRITY CHECKPOINT | 21 | 146 | 8.903e-27 | 1.883e-24 |
| 23 | ORGANELLE FISSION | 29 | 496 | 1.515e-25 | 3.065e-23 |
| 24 | POSITIVE REGULATION OF CELL CYCLE PROCESS | 23 | 247 | 8e-25 | 1.551e-22 |
| 25 | CELLULAR RESPONSE TO DNA DAMAGE STIMULUS | 30 | 720 | 3.486e-22 | 6.488e-20 |
| 26 | REGULATION OF CELL CYCLE G1 S PHASE TRANSITION | 18 | 147 | 1.067e-21 | 1.91e-19 |
| 27 | NEGATIVE REGULATION OF CELL CYCLE G1 S PHASE TRANSITION | 16 | 98 | 1.833e-21 | 3.16e-19 |
| 28 | NEGATIVE REGULATION OF PROTEIN METABOLIC PROCESS | 34 | 1087 | 2.55e-21 | 4.238e-19 |
| 29 | CHROMOSOME ORGANIZATION | 33 | 1009 | 3.04e-21 | 4.877e-19 |
| 30 | POSITIVE REGULATION OF PROTEIN METABOLIC PROCESS | 38 | 1492 | 6.154e-21 | 9.545e-19 |
| 31 | CELL CYCLE G2 M PHASE TRANSITION | 17 | 138 | 1.419e-20 | 2.13e-18 |
| 32 | NEGATIVE REGULATION OF PROTEIN MODIFICATION PROCESS | 27 | 616 | 1.829e-20 | 2.659e-18 |
| 33 | REGULATION OF ORGANELLE ORGANIZATION | 34 | 1178 | 3.22e-20 | 4.541e-18 |
| 34 | G1 DNA DAMAGE CHECKPOINT | 14 | 73 | 6.172e-20 | 8.446e-18 |
| 35 | SIGNAL TRANSDUCTION IN RESPONSE TO DNA DAMAGE | 15 | 96 | 7.228e-20 | 9.609e-18 |
| 36 | REGULATION OF PHOSPHORUS METABOLIC PROCESS | 38 | 1618 | 1.018e-19 | 1.316e-17 |
| 37 | POSITIVE REGULATION OF PROTEIN MODIFICATION PROCESS | 33 | 1135 | 1.121e-19 | 1.41e-17 |
| 38 | MITOTIC DNA INTEGRITY CHECKPOINT | 15 | 100 | 1.381e-19 | 1.692e-17 |
| 39 | REGULATION OF CELL DIVISION | 20 | 272 | 1.479e-19 | 1.765e-17 |
| 40 | REGULATION OF CELL CYCLE ARREST | 15 | 108 | 4.651e-19 | 5.41e-17 |
| 41 | REGULATION OF KINASE ACTIVITY | 28 | 776 | 5.34e-19 | 6.06e-17 |
| 42 | NEGATIVE REGULATION OF TRANSFERASE ACTIVITY | 21 | 351 | 1.163e-18 | 1.288e-16 |
| 43 | DNA METABOLIC PROCESS | 27 | 758 | 3.665e-18 | 3.965e-16 |
| 44 | POSITIVE REGULATION OF TRANSFERASE ACTIVITY | 25 | 616 | 3.915e-18 | 4.14e-16 |
| 45 | REGULATION OF CYCLIN DEPENDENT PROTEIN KINASE ACTIVITY | 14 | 97 | 4.31e-18 | 4.456e-16 |
| 46 | REGULATION OF LIGASE ACTIVITY | 15 | 130 | 8.35e-18 | 8.446e-16 |
| 47 | REGULATION OF NUCLEAR DIVISION | 16 | 163 | 8.675e-18 | 8.589e-16 |
| 48 | CELLULAR RESPONSE TO STRESS | 35 | 1565 | 2.458e-17 | 2.382e-15 |
| 49 | POSITIVE REGULATION OF CELL CYCLE ARREST | 13 | 85 | 3.272e-17 | 3.107e-15 |
| 50 | POSITIVE REGULATION OF PROTEOLYSIS | 20 | 363 | 4.286e-17 | 3.988e-15 |
| 51 | REGULATION OF CELL PROLIFERATION | 34 | 1496 | 5.202e-17 | 4.746e-15 |
| 52 | REGULATION OF SISTER CHROMATID SEGREGATION | 12 | 67 | 7.687e-17 | 6.879e-15 |
| 53 | REGULATION OF CELLULAR PROTEIN CATABOLIC PROCESS | 18 | 274 | 8.76e-17 | 7.69e-15 |
| 54 | REGULATION OF PROTEIN CATABOLIC PROCESS | 20 | 393 | 1.989e-16 | 1.714e-14 |
| 55 | NEGATIVE REGULATION OF CATALYTIC ACTIVITY | 26 | 829 | 3.815e-16 | 3.227e-14 |
| 56 | REGULATION OF PROTEIN SERINE THREONINE KINASE ACTIVITY | 21 | 470 | 4.319e-16 | 3.588e-14 |
| 57 | ANAPHASE PROMOTING COMPLEX DEPENDENT CATABOLIC PROCESS | 12 | 77 | 4.554e-16 | 3.718e-14 |
| 58 | POSITIVE REGULATION OF GENE EXPRESSION | 35 | 1733 | 5.813e-16 | 4.663e-14 |
| 59 | SISTER CHROMATID SEGREGATION | 15 | 176 | 8.459e-16 | 6.671e-14 |
| 60 | REGULATION OF PROTEOLYSIS | 24 | 711 | 1.313e-15 | 1.018e-13 |
| 61 | CHROMOSOME SEGREGATION | 17 | 272 | 1.576e-15 | 1.191e-13 |
| 62 | REGULATION OF CHROMOSOME SEGREGATION | 12 | 85 | 1.587e-15 | 1.191e-13 |
| 63 | NUCLEAR CHROMOSOME SEGREGATION | 16 | 228 | 1.88e-15 | 1.389e-13 |
| 64 | POSITIVE REGULATION OF BIOSYNTHETIC PROCESS | 35 | 1805 | 2.024e-15 | 1.472e-13 |
| 65 | REGULATION OF CHROMOSOME ORGANIZATION | 17 | 278 | 2.265e-15 | 1.621e-13 |
| 66 | NEGATIVE REGULATION OF MOLECULAR FUNCTION | 28 | 1079 | 2.62e-15 | 1.847e-13 |
| 67 | DNA REPLICATION INITIATION | 9 | 29 | 2.734e-15 | 1.899e-13 |
| 68 | POSITIVE REGULATION OF CELLULAR PROTEIN CATABOLIC PROCESS | 15 | 192 | 3.118e-15 | 2.134e-13 |
| 69 | POSITIVE REGULATION OF MITOTIC CELL CYCLE | 13 | 123 | 4.787e-15 | 3.228e-13 |
| 70 | SIGNAL TRANSDUCTION BY P53 CLASS MEDIATOR | 13 | 127 | 7.314e-15 | 4.862e-13 |
| 71 | DNA REPLICATION | 15 | 208 | 1.027e-14 | 6.731e-13 |
| 72 | REGULATION OF PROTEIN UBIQUITINATION INVOLVED IN UBIQUITIN DEPENDENT PROTEIN CATABOLIC PROCESS | 12 | 103 | 1.735e-14 | 1.121e-12 |
| 73 | POSITIVE REGULATION OF CELL PROLIFERATION | 24 | 814 | 2.597e-14 | 1.655e-12 |
| 74 | POSITIVE REGULATION OF CATALYTIC ACTIVITY | 31 | 1518 | 3.81e-14 | 2.395e-12 |
| 75 | POSITIVE REGULATION OF LIGASE ACTIVITY | 12 | 110 | 3.898e-14 | 2.418e-12 |
| 76 | SISTER CHROMATID COHESION | 12 | 111 | 4.356e-14 | 2.667e-12 |
| 77 | REGULATION OF PROTEIN MODIFICATION BY SMALL PROTEIN CONJUGATION OR REMOVAL | 16 | 280 | 4.698e-14 | 2.839e-12 |
| 78 | POSITIVE REGULATION OF CELL DEATH | 21 | 605 | 6.298e-14 | 3.757e-12 |
| 79 | CELL CYCLE ARREST | 13 | 154 | 9.18e-14 | 5.407e-12 |
| 80 | POSITIVE REGULATION OF PROTEIN MODIFICATION BY SMALL PROTEIN CONJUGATION OR REMOVAL | 14 | 196 | 9.754e-14 | 5.673e-12 |
| 81 | POSITIVE REGULATION OF CELLULAR PROTEIN LOCALIZATION | 17 | 360 | 1.576e-13 | 9.055e-12 |
| 82 | NEGATIVE REGULATION OF CHROMOSOME SEGREGATION | 8 | 28 | 2.203e-13 | 1.25e-11 |
| 83 | REGULATION OF CATABOLIC PROCESS | 22 | 731 | 2.556e-13 | 1.433e-11 |
| 84 | POSITIVE REGULATION OF PROTEIN CATABOLIC PROCESS | 15 | 263 | 3.226e-13 | 1.787e-11 |
| 85 | PROTEIN UBIQUITINATION INVOLVED IN UBIQUITIN DEPENDENT PROTEIN CATABOLIC PROCESS | 12 | 134 | 4.305e-13 | 2.357e-11 |
| 86 | POSITIVE REGULATION OF MOLECULAR FUNCTION | 32 | 1791 | 5.034e-13 | 2.723e-11 |
| 87 | PROTEIN PHOSPHORYLATION | 24 | 944 | 6.417e-13 | 3.432e-11 |
| 88 | NEGATIVE REGULATION OF PROTEIN MODIFICATION BY SMALL PROTEIN CONJUGATION OR REMOVAL | 12 | 139 | 6.694e-13 | 3.539e-11 |
| 89 | POSITIVE REGULATION OF FIBROBLAST PROLIFERATION | 9 | 53 | 1.114e-12 | 5.825e-11 |
| 90 | NEGATIVE REGULATION OF MITOTIC NUCLEAR DIVISION | 8 | 34 | 1.261e-12 | 6.517e-11 |
| 91 | REGULATION OF CELLULAR PROTEIN LOCALIZATION | 19 | 552 | 1.376e-12 | 7.034e-11 |
| 92 | POSITIVE REGULATION OF MITOCHONDRIAL OUTER MEMBRANE PERMEABILIZATION INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 8 | 36 | 2.087e-12 | 1.055e-10 |
| 93 | NEGATIVE REGULATION OF CELL DIVISION | 9 | 60 | 3.628e-12 | 1.796e-10 |
| 94 | RESPONSE TO ABIOTIC STIMULUS | 24 | 1024 | 3.622e-12 | 1.796e-10 |
| 95 | REGULATION OF CELL DEATH | 28 | 1472 | 5.467e-12 | 2.677e-10 |
| 96 | PROTEASOMAL PROTEIN CATABOLIC PROCESS | 14 | 271 | 8.098e-12 | 3.925e-10 |
| 97 | CELLULAR RESPONSE TO UV | 9 | 66 | 8.9e-12 | 4.269e-10 |
| 98 | REGULATION OF MITOCHONDRIAL OUTER MEMBRANE PERMEABILIZATION INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 8 | 43 | 9.762e-12 | 4.635e-10 |
| 99 | POSITIVE REGULATION OF CELL CYCLE PHASE TRANSITION | 9 | 68 | 1.177e-11 | 5.531e-10 |
| 100 | DNA DEPENDENT DNA REPLICATION | 10 | 99 | 1.277e-11 | 5.943e-10 |
| 101 | DNA REPAIR | 17 | 480 | 1.561e-11 | 7.193e-10 |
| 102 | REGULATION OF TRANSCRIPTION INVOLVED IN G1 S TRANSITION OF MITOTIC CELL CYCLE | 7 | 27 | 1.613e-11 | 7.36e-10 |
| 103 | REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 30 | 1784 | 1.652e-11 | 7.465e-10 |
| 104 | NEGATIVE REGULATION OF NUCLEAR DIVISION | 8 | 46 | 1.739e-11 | 7.778e-10 |
| 105 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 23 | 1004 | 1.798e-11 | 7.966e-10 |
| 106 | REGULATION OF PROTEIN INSERTION INTO MITOCHONDRIAL MEMBRANE INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 7 | 29 | 2.817e-11 | 1.225e-09 |
| 107 | POSITIVE REGULATION OF PROTEIN INSERTION INTO MITOCHONDRIAL MEMBRANE INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 7 | 29 | 2.817e-11 | 1.225e-09 |
| 108 | REGULATION OF PROTEASOMAL UBIQUITIN DEPENDENT PROTEIN CATABOLIC PROCESS | 11 | 148 | 3.341e-11 | 1.426e-09 |
| 109 | PEPTIDYL AMINO ACID MODIFICATION | 21 | 841 | 3.335e-11 | 1.426e-09 |
| 110 | REGULATION OF FIBROBLAST PROLIFERATION | 9 | 81 | 5.958e-11 | 2.52e-09 |
| 111 | PROTEIN MODIFICATION BY SMALL PROTEIN CONJUGATION OR REMOVAL | 21 | 873 | 6.662e-11 | 2.793e-09 |
| 112 | NEGATIVE REGULATION OF PHOSPHORUS METABOLIC PROCESS | 17 | 541 | 1.009e-10 | 4.157e-09 |
| 113 | NEGATIVE REGULATION OF PHOSPHATE METABOLIC PROCESS | 17 | 541 | 1.009e-10 | 4.157e-09 |
| 114 | POSITIVE REGULATION OF CATABOLIC PROCESS | 15 | 395 | 1.065e-10 | 4.349e-09 |
| 115 | RESPONSE TO UV | 10 | 126 | 1.44e-10 | 5.826e-09 |
| 116 | PHOSPHORYLATION | 24 | 1228 | 1.582e-10 | 6.348e-09 |
| 117 | REGULATION OF DNA METABOLIC PROCESS | 14 | 340 | 1.65e-10 | 6.563e-09 |
| 118 | CELLULAR RESPONSE TO LIGHT STIMULUS | 9 | 91 | 1.73e-10 | 6.763e-09 |
| 119 | MITOTIC SISTER CHROMATID SEGREGATION | 9 | 91 | 1.73e-10 | 6.763e-09 |
| 120 | NEGATIVE REGULATION OF PHOSPHORYLATION | 15 | 422 | 2.671e-10 | 1.036e-08 |
| 121 | REGULATION OF PROTEASOMAL PROTEIN CATABOLIC PROCESS | 11 | 181 | 2.934e-10 | 1.128e-08 |
| 122 | CELLULAR RESPONSE TO RADIATION | 10 | 137 | 3.301e-10 | 1.259e-08 |
| 123 | CELL PROLIFERATION | 18 | 672 | 3.501e-10 | 1.324e-08 |
| 124 | MEIOTIC CELL CYCLE | 11 | 186 | 3.925e-10 | 1.473e-08 |
| 125 | POSITIVE REGULATION OF INTRACELLULAR TRANSPORT | 14 | 370 | 4.985e-10 | 1.855e-08 |
| 126 | REGULATION OF MEMBRANE PERMEABILITY | 8 | 70 | 5.793e-10 | 2.139e-08 |
| 127 | POSITIVE REGULATION OF RESPONSE TO STIMULUS | 29 | 1929 | 5.918e-10 | 2.168e-08 |
| 128 | RESPONSE TO OXYGEN LEVELS | 13 | 311 | 6.559e-10 | 2.366e-08 |
| 129 | RESPONSE TO LIPID | 20 | 888 | 6.531e-10 | 2.366e-08 |
| 130 | CELL DEATH | 21 | 1001 | 8.095e-10 | 2.897e-08 |
| 131 | SPINDLE CHECKPOINT | 6 | 25 | 8.176e-10 | 2.904e-08 |
| 132 | INTRACELLULAR SIGNAL TRANSDUCTION | 26 | 1572 | 8.312e-10 | 2.93e-08 |
| 133 | NEGATIVE REGULATION OF PROTEIN CATABOLIC PROCESS | 9 | 109 | 8.846e-10 | 3.095e-08 |
| 134 | CELLULAR RESPONSE TO ENDOGENOUS STIMULUS | 21 | 1008 | 9.177e-10 | 3.187e-08 |
| 135 | POSITIVE REGULATION OF PHOSPHATE METABOLIC PROCESS | 21 | 1036 | 1.501e-09 | 5.134e-08 |
| 136 | POSITIVE REGULATION OF PHOSPHORUS METABOLIC PROCESS | 21 | 1036 | 1.501e-09 | 5.134e-08 |
| 137 | POSITIVE REGULATION OF STEM CELL DIFFERENTIATION | 7 | 50 | 1.678e-09 | 5.698e-08 |
| 138 | REGULATION OF PROTEIN LOCALIZATION | 20 | 950 | 2.093e-09 | 7.056e-08 |
| 139 | POSITIVE REGULATION OF ORGANELLE ORGANIZATION | 16 | 573 | 2.116e-09 | 7.084e-08 |
| 140 | POSITIVE REGULATION OF CELL COMMUNICATION | 25 | 1532 | 2.56e-09 | 8.509e-08 |
| 141 | POSITIVE REGULATION OF APOPTOTIC SIGNALING PATHWAY | 10 | 171 | 2.887e-09 | 9.526e-08 |
| 142 | RESPONSE TO DRUG | 14 | 431 | 3.564e-09 | 1.167e-07 |
| 143 | NEGATIVE REGULATION OF CELLULAR COMPONENT ORGANIZATION | 17 | 684 | 3.586e-09 | 1.167e-07 |
| 144 | POSITIVE REGULATION OF ESTABLISHMENT OF PROTEIN LOCALIZATION | 15 | 514 | 3.962e-09 | 1.28e-07 |
| 145 | REGULATION OF CELLULAR RESPONSE TO STRESS | 17 | 691 | 4.175e-09 | 1.34e-07 |
| 146 | RESPONSE TO ENDOGENOUS STIMULUS | 24 | 1450 | 4.434e-09 | 1.413e-07 |
| 147 | MITOCHONDRIAL MEMBRANE ORGANIZATION | 8 | 92 | 5.297e-09 | 1.677e-07 |
| 148 | POSITIVE REGULATION OF EPITHELIAL TO MESENCHYMAL TRANSITION | 6 | 34 | 6.022e-09 | 1.893e-07 |
| 149 | REGULATION OF INTRACELLULAR TRANSPORT | 16 | 621 | 6.644e-09 | 2.075e-07 |
| 150 | RESPONSE TO ORGANIC CYCLIC COMPOUND | 19 | 917 | 7.394e-09 | 2.293e-07 |
| 151 | NEGATIVE REGULATION OF CHROMOSOME ORGANIZATION | 8 | 96 | 7.443e-09 | 2.293e-07 |
| 152 | POSITIVE REGULATION OF CYCLIN DEPENDENT PROTEIN KINASE ACTIVITY | 6 | 36 | 8.663e-09 | 2.652e-07 |
| 153 | NEGATIVE REGULATION OF KINASE ACTIVITY | 11 | 250 | 8.824e-09 | 2.683e-07 |
| 154 | NEGATIVE REGULATION OF ORGANELLE ORGANIZATION | 13 | 387 | 9.151e-09 | 2.765e-07 |
| 155 | POSITIVE REGULATION OF CELLULAR COMPONENT ORGANIZATION | 21 | 1152 | 9.817e-09 | 2.947e-07 |
| 156 | NEGATIVE REGULATION OF CELLULAR PROTEIN CATABOLIC PROCESS | 7 | 64 | 9.946e-09 | 2.967e-07 |
| 157 | NEGATIVE REGULATION OF CELL PROLIFERATION | 16 | 643 | 1.085e-08 | 3.216e-07 |
| 158 | DIGESTIVE SYSTEM DEVELOPMENT | 9 | 148 | 1.33e-08 | 3.916e-07 |
| 159 | CELLULAR RESPONSE TO ABIOTIC STIMULUS | 11 | 263 | 1.491e-08 | 4.362e-07 |
| 160 | MEIOTIC CELL CYCLE PROCESS | 9 | 152 | 1.678e-08 | 4.881e-07 |
| 161 | PROTEIN CATABOLIC PROCESS | 15 | 579 | 1.946e-08 | 5.626e-07 |
| 162 | RESPONSE TO RADIATION | 13 | 413 | 1.977e-08 | 5.678e-07 |
| 163 | RESPONSE TO STEROID HORMONE | 14 | 497 | 2.158e-08 | 6.161e-07 |
| 164 | PHOSPHATE CONTAINING COMPOUND METABOLIC PROCESS | 27 | 1977 | 2.256e-08 | 6.401e-07 |
| 165 | REGULATION OF DNA REPLICATION | 9 | 161 | 2.769e-08 | 7.808e-07 |
| 166 | PROTEIN SUMOYLATION | 8 | 115 | 3.11e-08 | 8.719e-07 |
| 167 | NEGATIVE REGULATION OF NITROGEN COMPOUND METABOLIC PROCESS | 23 | 1517 | 5.191e-08 | 1.446e-06 |
| 168 | PROTEIN UBIQUITINATION | 15 | 629 | 5.786e-08 | 1.603e-06 |
| 169 | POSITIVE REGULATION OF CHROMOSOME SEGREGATION | 5 | 25 | 6.116e-08 | 1.684e-06 |
| 170 | NEGATIVE REGULATION OF PROTEIN SERINE THREONINE KINASE ACTIVITY | 8 | 126 | 6.36e-08 | 1.741e-06 |
| 171 | REGULATION OF ESTABLISHMENT OF PROTEIN LOCALIZATION TO MITOCHONDRION | 8 | 128 | 7.191e-08 | 1.957e-06 |
| 172 | CELLULAR RESPONSE TO ORGANIC CYCLIC COMPOUND | 13 | 465 | 7.912e-08 | 2.14e-06 |
| 173 | REGULATION OF MICROTUBULE BASED PROCESS | 10 | 243 | 8.209e-08 | 2.208e-06 |
| 174 | RESPONSE TO GROWTH FACTOR | 13 | 475 | 1.012e-07 | 2.706e-06 |
| 175 | POSITIVE REGULATION OF KINASE ACTIVITY | 13 | 482 | 1.198e-07 | 3.186e-06 |
| 176 | NEGATIVE REGULATION OF DNA REPLICATION | 6 | 55 | 1.209e-07 | 3.196e-06 |
| 177 | NEGATIVE REGULATION OF PROTEOLYSIS | 11 | 329 | 1.458e-07 | 3.83e-06 |
| 178 | REPLICATIVE SENESCENCE | 4 | 12 | 1.465e-07 | 3.83e-06 |
| 179 | RESPONSE TO HORMONE | 17 | 893 | 1.751e-07 | 4.551e-06 |
| 180 | REGULATION OF CELL CYCLE G2 M PHASE TRANSITION | 6 | 59 | 1.854e-07 | 4.792e-06 |
| 181 | NEGATIVE REGULATION OF CATABOLIC PROCESS | 9 | 203 | 2.025e-07 | 5.206e-06 |
| 182 | POSITIVE REGULATION OF TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 7 | 100 | 2.266e-07 | 5.794e-06 |
| 183 | REGULATION OF TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 9 | 207 | 2.389e-07 | 6.075e-06 |
| 184 | G2 DNA DAMAGE CHECKPOINT | 5 | 33 | 2.66e-07 | 6.691e-06 |
| 185 | REGULATION OF CELL AGING | 5 | 33 | 2.66e-07 | 6.691e-06 |
| 186 | REGULATION OF CELLULAR LOCALIZATION | 20 | 1277 | 2.833e-07 | 7.087e-06 |
| 187 | CELLULAR RESPONSE TO REACTIVE OXYGEN SPECIES | 7 | 104 | 2.967e-07 | 7.382e-06 |
| 188 | RESPONSE TO LIGHT STIMULUS | 10 | 280 | 3.059e-07 | 7.571e-06 |
| 189 | REGULATION OF BINDING | 10 | 283 | 3.374e-07 | 8.307e-06 |
| 190 | REPRODUCTION | 20 | 1297 | 3.63e-07 | 8.889e-06 |
| 191 | REGULATION OF MITOCHONDRION ORGANIZATION | 9 | 218 | 3.7e-07 | 8.966e-06 |
| 192 | REGULATION OF PROTEIN LOCALIZATION TO NUCLEUS | 9 | 218 | 3.7e-07 | 8.966e-06 |
| 193 | ACTIVATION OF ANAPHASE PROMOTING COMPLEX ACTIVITY | 4 | 15 | 4.002e-07 | 9.598e-06 |
| 194 | REGULATION OF EPITHELIAL TO MESENCHYMAL TRANSITION | 6 | 67 | 3.996e-07 | 9.598e-06 |
| 195 | POSITIVE REGULATION OF CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 8 | 162 | 4.418e-07 | 1.054e-05 |
| 196 | CELLULAR RESPONSE TO ORGANIC SUBSTANCE | 24 | 1848 | 4.448e-07 | 1.056e-05 |
| 197 | REGULATION OF STEM CELL DIFFERENTIATION | 7 | 113 | 5.23e-07 | 1.235e-05 |
| 198 | RHYTHMIC PROCESS | 10 | 298 | 5.416e-07 | 1.273e-05 |
| 199 | MESENCHYME MORPHOGENESIS | 5 | 38 | 5.533e-07 | 1.294e-05 |
| 200 | POSITIVE REGULATION OF MITOCHONDRION ORGANIZATION | 8 | 167 | 5.568e-07 | 1.295e-05 |
| 201 | NEGATIVE REGULATION OF CELL AGING | 4 | 17 | 6.932e-07 | 1.605e-05 |
| 202 | REGULATION OF DNA DEPENDENT DNA REPLICATION | 5 | 41 | 8.179e-07 | 1.884e-05 |
| 203 | PEPTIDYL LYSINE MODIFICATION | 10 | 312 | 8.226e-07 | 1.886e-05 |
| 204 | MITOCHONDRIAL TRANSPORT | 8 | 177 | 8.652e-07 | 1.973e-05 |
| 205 | GLAND DEVELOPMENT | 11 | 395 | 8.941e-07 | 2.029e-05 |
| 206 | REGULATION OF MICROTUBULE POLYMERIZATION OR DEPOLYMERIZATION | 8 | 178 | 9.028e-07 | 2.039e-05 |
| 207 | POSITIVE REGULATION OF PROTEIN LOCALIZATION TO NUCLEUS | 7 | 129 | 1.282e-06 | 2.881e-05 |
| 208 | PROTEIN LOCALIZATION TO CHROMOSOME | 5 | 45 | 1.316e-06 | 2.943e-05 |
| 209 | REGULATION OF CYTOSKELETON ORGANIZATION | 12 | 502 | 1.368e-06 | 3.046e-05 |
| 210 | ORGAN REGENERATION | 6 | 83 | 1.432e-06 | 3.172e-05 |
| 211 | CELLULAR RESPONSE TO NITROGEN COMPOUND | 12 | 505 | 1.456e-06 | 3.21e-05 |
| 212 | POSITIVE REGULATION OF CELL DIVISION | 7 | 132 | 1.496e-06 | 3.283e-05 |
| 213 | REGULATION OF CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 10 | 337 | 1.65e-06 | 3.604e-05 |
| 214 | MEIOSIS I | 6 | 88 | 2.021e-06 | 4.394e-05 |
| 215 | MICROTUBULE CYTOSKELETON ORGANIZATION | 10 | 348 | 2.2e-06 | 4.761e-05 |
| 216 | PROTEOLYSIS | 18 | 1208 | 2.55e-06 | 5.493e-05 |
| 217 | NEGATIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 14 | 740 | 2.655e-06 | 5.692e-05 |
| 218 | REGULATION OF RESPONSE TO DNA DAMAGE STIMULUS | 7 | 145 | 2.803e-06 | 5.982e-05 |
| 219 | CHROMATIN MODIFICATION | 12 | 539 | 2.857e-06 | 6.069e-05 |
| 220 | RESPONSE TO NITROGEN COMPOUND | 15 | 859 | 2.991e-06 | 6.327e-05 |
| 221 | RESPONSE TO ALCOHOL | 10 | 362 | 3.126e-06 | 6.582e-05 |
| 222 | PEPTIDYL SERINE MODIFICATION | 7 | 148 | 3.211e-06 | 6.7e-05 |
| 223 | REGULATION OF APOPTOTIC SIGNALING PATHWAY | 10 | 363 | 3.204e-06 | 6.7e-05 |
| 224 | NEGATIVE REGULATION OF GENE EXPRESSION | 20 | 1493 | 3.249e-06 | 6.749e-05 |
| 225 | POSITIVE REGULATION OF CHROMOSOME ORGANIZATION | 7 | 150 | 3.51e-06 | 7.26e-05 |
| 226 | NEGATIVE REGULATION OF CELL DEATH | 15 | 872 | 3.593e-06 | 7.397e-05 |
| 227 | CELLULAR RESPONSE TO LIPID | 11 | 457 | 3.664e-06 | 7.51e-05 |
| 228 | APOPTOTIC SIGNALING PATHWAY | 9 | 289 | 3.822e-06 | 7.753e-05 |
| 229 | POSITIVE REGULATION OF PROTEIN SERINE THREONINE KINASE ACTIVITY | 9 | 289 | 3.822e-06 | 7.753e-05 |
| 230 | INTRINSIC APOPTOTIC SIGNALING PATHWAY | 7 | 152 | 3.833e-06 | 7.753e-05 |
| 231 | SMAD PROTEIN SIGNAL TRANSDUCTION | 5 | 56 | 3.966e-06 | 7.988e-05 |
| 232 | REGULATION OF TRANSFORMING GROWTH FACTOR BETA RECEPTOR SIGNALING PATHWAY | 6 | 99 | 4.026e-06 | 8.04e-05 |
| 233 | REGULATION OF CELLULAR RESPONSE TO TRANSFORMING GROWTH FACTOR BETA STIMULUS | 6 | 99 | 4.026e-06 | 8.04e-05 |
| 234 | RESPONSE TO ESTROGEN | 8 | 218 | 4.104e-06 | 8.131e-05 |
| 235 | RESPONSE TO OXYGEN CONTAINING COMPOUND | 19 | 1381 | 4.107e-06 | 8.131e-05 |
| 236 | REGULATION OF CELLULAR SENESCENCE | 4 | 26 | 4.229e-06 | 8.338e-05 |
| 237 | CHROMATIN ORGANIZATION | 13 | 663 | 4.273e-06 | 8.389e-05 |
| 238 | REGULATION OF NUCLEOCYTOPLASMIC TRANSPORT | 8 | 220 | 4.39e-06 | 8.582e-05 |
| 239 | NEGATIVE REGULATION OF CELLULAR CATABOLIC PROCESS | 7 | 156 | 4.551e-06 | 8.86e-05 |
| 240 | PROTEIN K11 LINKED UBIQUITINATION | 4 | 27 | 4.949e-06 | 9.594e-05 |
| 241 | POSITIVE REGULATION OF DEVELOPMENTAL PROCESS | 17 | 1142 | 5.163e-06 | 9.969e-05 |
| 242 | REGENERATION | 7 | 161 | 5.604e-06 | 0.0001077 |
| 243 | RESPONSE TO INORGANIC SUBSTANCE | 11 | 479 | 5.731e-06 | 0.0001097 |
| 244 | REGULATION OF SIGNAL TRANSDUCTION BY P53 CLASS MEDIATOR | 7 | 162 | 5.837e-06 | 0.0001113 |
| 245 | CELLULAR RESPONSE TO HYDROGEN PEROXIDE | 5 | 61 | 6.074e-06 | 0.0001154 |
| 246 | MACROMOLECULE CATABOLIC PROCESS | 15 | 926 | 7.424e-06 | 0.0001404 |
| 247 | NEGATIVE REGULATION OF DNA METABOLIC PROCESS | 6 | 111 | 7.812e-06 | 0.0001472 |
| 248 | POSITIVE REGULATION OF TRANSPORT | 15 | 936 | 8.442e-06 | 0.0001584 |
| 249 | GROWTH | 10 | 410 | 9.346e-06 | 0.0001746 |
| 250 | REGULATION OF RESPONSE TO STRESS | 19 | 1468 | 9.898e-06 | 0.0001842 |
| 251 | TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 13 | 724 | 1.101e-05 | 0.000204 |
| 252 | MITOTIC SPINDLE ORGANIZATION | 5 | 69 | 1.117e-05 | 0.0002063 |
| 253 | NEGATIVE REGULATION OF CELLULAR SENESCENCE | 3 | 11 | 1.193e-05 | 0.0002185 |
| 254 | ACTIVATION OF MAPKKK ACTIVITY | 3 | 11 | 1.193e-05 | 0.0002185 |
| 255 | RESPONSE TO KETONE | 7 | 182 | 1.25e-05 | 0.0002271 |
| 256 | REGULATION OF CELL DIFFERENTIATION | 19 | 1492 | 1.246e-05 | 0.0002271 |
| 257 | INTRINSIC APOPTOTIC SIGNALING PATHWAY IN RESPONSE TO DNA DAMAGE | 5 | 71 | 1.286e-05 | 0.0002301 |
| 258 | MICROTUBULE BASED PROCESS | 11 | 522 | 1.285e-05 | 0.0002301 |
| 259 | PROTEIN DESTABILIZATION | 4 | 34 | 1.278e-05 | 0.0002301 |
| 260 | POSITIVE REGULATION OF NUCLEOCYTOPLASMIC TRANSPORT | 6 | 121 | 1.282e-05 | 0.0002301 |
| 261 | CELLULAR RESPONSE TO OXIDATIVE STRESS | 7 | 184 | 1.342e-05 | 0.0002392 |
| 262 | POSITIVE REGULATION OF DNA METABOLIC PROCESS | 7 | 185 | 1.39e-05 | 0.0002468 |
| 263 | RESPONSE TO MINERALOCORTICOID | 4 | 35 | 1.439e-05 | 0.0002536 |
| 264 | RESPONSE TO IRON ION | 4 | 35 | 1.439e-05 | 0.0002536 |
| 265 | MITOTIC SISTER CHROMATID COHESION | 3 | 12 | 1.585e-05 | 0.0002784 |
| 266 | CELLULAR RESPONSE TO EXTERNAL STIMULUS | 8 | 264 | 1.659e-05 | 0.0002902 |
| 267 | RESPONSE TO REACTIVE OXYGEN SPECIES | 7 | 191 | 1.708e-05 | 0.0002977 |
| 268 | REGULATION OF CELLULAR RESPONSE TO HEAT | 5 | 76 | 1.794e-05 | 0.0003116 |
| 269 | MITOTIC CELL CYCLE ARREST | 3 | 13 | 2.055e-05 | 0.0003554 |
| 270 | CELLULAR RESPONSE TO HORMONE STIMULUS | 11 | 552 | 2.157e-05 | 0.0003703 |
| 271 | REGULATION OF CELL MORPHOGENESIS | 11 | 552 | 2.157e-05 | 0.0003703 |
| 272 | DNA GEOMETRIC CHANGE | 5 | 81 | 2.448e-05 | 0.0004187 |
| 273 | POSITIVE REGULATION OF MITOTIC SISTER CHROMATID SEPARATION | 3 | 14 | 2.607e-05 | 0.0004395 |
| 274 | POSITIVE REGULATION OF MITOTIC METAPHASE ANAPHASE TRANSITION | 3 | 14 | 2.607e-05 | 0.0004395 |
| 275 | POSITIVE REGULATION OF P38MAPK CASCADE | 3 | 14 | 2.607e-05 | 0.0004395 |
| 276 | POSITIVE REGULATION OF METAPHASE ANAPHASE TRANSITION OF CELL CYCLE | 3 | 14 | 2.607e-05 | 0.0004395 |
| 277 | REGULATION OF EPITHELIAL CELL PROLIFERATION | 8 | 285 | 2.87e-05 | 0.0004822 |
| 278 | NEGATIVE REGULATION OF DEVELOPMENTAL PROCESS | 13 | 801 | 3.181e-05 | 0.0005324 |
| 279 | POSITIVE REGULATION OF DNA REPLICATION | 5 | 86 | 3.272e-05 | 0.0005457 |
| 280 | CELLULAR RESPONSE TO OXYGEN LEVELS | 6 | 143 | 3.307e-05 | 0.0005495 |
| 281 | RESPONSE TO TRANSFORMING GROWTH FACTOR BETA | 6 | 144 | 3.439e-05 | 0.0005694 |
| 282 | RESPONSE TO IONIZING RADIATION | 6 | 145 | 3.575e-05 | 0.0005899 |
| 283 | REGULATION OF PROTEIN COMPLEX DISASSEMBLY | 7 | 217 | 3.872e-05 | 0.0006366 |
| 284 | NEGATIVE REGULATION OF DNA DEPENDENT DNA REPLICATION | 3 | 16 | 3.986e-05 | 0.0006509 |
| 285 | CELLULAR RESPONSE TO STEROID HORMONE STIMULUS | 7 | 218 | 3.987e-05 | 0.0006509 |
| 286 | EPITHELIUM DEVELOPMENT | 14 | 945 | 4.166e-05 | 0.0006777 |
| 287 | REGULATION OF PROTEIN STABILITY | 7 | 221 | 4.348e-05 | 0.0007049 |
| 288 | REGULATION OF SISTER CHROMATID COHESION | 3 | 17 | 4.826e-05 | 0.0007796 |
| 289 | POSITIVE REGULATION OF PATHWAY RESTRICTED SMAD PROTEIN PHOSPHORYLATION | 4 | 48 | 5.126e-05 | 0.0008253 |
| 290 | TRANSFORMING GROWTH FACTOR BETA RECEPTOR SIGNALING PATHWAY | 5 | 95 | 5.283e-05 | 0.0008477 |
| 291 | REGULATION OF CELLULAR RESPONSE TO GROWTH FACTOR STIMULUS | 7 | 229 | 5.445e-05 | 0.0008707 |
| 292 | TISSUE DEVELOPMENT | 18 | 1518 | 5.67e-05 | 0.0009036 |
| 293 | POSITIVE REGULATION OF CELL CYCLE G2 M PHASE TRANSITION | 3 | 18 | 5.773e-05 | 0.0009137 |
| 294 | REGULATION OF UBIQUITIN PROTEIN LIGASE ACTIVITY | 3 | 18 | 5.773e-05 | 0.0009137 |
| 295 | POSITIVE REGULATION OF PROTEASOMAL PROTEIN CATABOLIC PROCESS | 5 | 98 | 6.131e-05 | 0.000967 |
| 296 | RESPONSE TO AMMONIUM ION | 4 | 51 | 6.52e-05 | 0.001025 |
| 297 | INTRINSIC APOPTOTIC SIGNALING PATHWAY BY P53 CLASS MEDIATOR | 4 | 53 | 7.591e-05 | 0.001189 |
| 298 | PROTEIN POLYUBIQUITINATION | 7 | 243 | 7.908e-05 | 0.001235 |
| 299 | NEGATIVE REGULATION OF CELL CYCLE ARREST | 3 | 20 | 8.016e-05 | 0.001247 |
| 300 | RESPONSE TO METAL ION | 8 | 333 | 8.581e-05 | 0.001327 |
| 301 | DEVELOPMENTAL GROWTH | 8 | 333 | 8.581e-05 | 0.001327 |
| 302 | NEGATIVE REGULATION OF PROTEIN COMPLEX DISASSEMBLY | 6 | 170 | 8.668e-05 | 0.001335 |
| 303 | RESPONSE TO HYDROGEN PEROXIDE | 5 | 109 | 0.0001017 | 0.001562 |
| 304 | ENDOCARDIAL CUSHION MORPHOGENESIS | 3 | 22 | 0.0001076 | 0.001647 |
| 305 | TUBE DEVELOPMENT | 10 | 552 | 0.0001155 | 0.001762 |
| 306 | POSITIVE REGULATION OF MULTICELLULAR ORGANISMAL METABOLIC PROCESS | 3 | 23 | 0.0001234 | 0.001855 |
| 307 | RESPONSE TO INCREASED OXYGEN LEVELS | 3 | 23 | 0.0001234 | 0.001855 |
| 308 | REGULATION OF PATHWAY RESTRICTED SMAD PROTEIN PHOSPHORYLATION | 4 | 60 | 0.0001236 | 0.001855 |
| 309 | RESPONSE TO HYPEROXIA | 3 | 23 | 0.0001234 | 0.001855 |
| 310 | POSITIVE REGULATION OF COLLAGEN METABOLIC PROCESS | 3 | 23 | 0.0001234 | 0.001855 |
| 311 | REGULATION OF PROTEIN IMPORT | 6 | 183 | 0.0001299 | 0.001944 |
| 312 | NEGATIVE REGULATION OF EPITHELIAL CELL PROLIFERATION | 5 | 115 | 0.000131 | 0.001953 |
| 313 | AGING | 7 | 264 | 0.0001324 | 0.001969 |
| 314 | CELLULAR RESPONSE TO OXYGEN CONTAINING COMPOUND | 12 | 799 | 0.0001374 | 0.002036 |
| 315 | POSITIVE REGULATION OF CELLULAR RESPONSE TO TRANSFORMING GROWTH FACTOR BETA STIMULUS | 3 | 24 | 0.0001406 | 0.002057 |
| 316 | POSITIVE REGULATION OF G1 S TRANSITION OF MITOTIC CELL CYCLE | 3 | 24 | 0.0001406 | 0.002057 |
| 317 | SOMITOGENESIS | 4 | 62 | 0.0001405 | 0.002057 |
| 318 | POSITIVE REGULATION OF TRANSFORMING GROWTH FACTOR BETA RECEPTOR SIGNALING PATHWAY | 3 | 24 | 0.0001406 | 0.002057 |
| 319 | REGULATION OF TRANSFORMING GROWTH FACTOR BETA PRODUCTION | 3 | 25 | 0.0001593 | 0.002289 |
| 320 | REGULATION OF PROTEIN ACETYLATION | 4 | 64 | 0.000159 | 0.002289 |
| 321 | EPITHELIAL CELL APOPTOTIC PROCESS | 3 | 25 | 0.0001593 | 0.002289 |
| 322 | MESENCHYME DEVELOPMENT | 6 | 190 | 0.0001594 | 0.002289 |
| 323 | POSITIVE REGULATION OF RESPONSE TO DNA DAMAGE STIMULUS | 4 | 64 | 0.000159 | 0.002289 |
| 324 | HISTONE PHOSPHORYLATION | 3 | 25 | 0.0001593 | 0.002289 |
| 325 | HEART DEVELOPMENT | 9 | 466 | 0.0001609 | 0.002303 |
| 326 | DNA CONFORMATION CHANGE | 7 | 273 | 0.0001628 | 0.002324 |
| 327 | DEVELOPMENTAL MATURATION | 6 | 193 | 0.0001736 | 0.00247 |
| 328 | IMMUNE SYSTEM DEVELOPMENT | 10 | 582 | 0.0001776 | 0.00252 |
| 329 | RESPONSE TO CORTICOSTERONE | 3 | 26 | 0.0001795 | 0.002523 |
| 330 | REGULATION OF P38MAPK CASCADE | 3 | 26 | 0.0001795 | 0.002523 |
| 331 | LENS DEVELOPMENT IN CAMERA TYPE EYE | 4 | 66 | 0.0001792 | 0.002523 |
| 332 | CELL AGING | 4 | 67 | 0.0001899 | 0.002662 |
| 333 | REGULATION OF MULTICELLULAR ORGANISMAL DEVELOPMENT | 18 | 1672 | 0.000194 | 0.002711 |
| 334 | POSITIVE REGULATION OF CYTOPLASMIC TRANSPORT | 7 | 282 | 0.0001987 | 0.002768 |
| 335 | PROTEIN COMPLEX SUBUNIT ORGANIZATION | 17 | 1527 | 0.000203 | 0.00282 |
| 336 | REGULATION OF CYTOPLASMIC TRANSPORT | 9 | 481 | 0.0002037 | 0.002821 |
| 337 | REGULATION OF CELL DEVELOPMENT | 12 | 836 | 0.000209 | 0.002886 |
| 338 | REGULATION OF INTRACELLULAR PROTEIN TRANSPORT | 8 | 381 | 0.0002161 | 0.002975 |
| 339 | ORGAN MORPHOGENESIS | 12 | 841 | 0.0002208 | 0.003031 |
| 340 | REGULATION OF DNA DAMAGE RESPONSE SIGNAL TRANSDUCTION BY P53 CLASS MEDIATOR | 3 | 28 | 0.0002248 | 0.003067 |
| 341 | NEGATIVE REGULATION OF DENDRITE DEVELOPMENT | 3 | 28 | 0.0002248 | 0.003067 |
| 342 | POSITIVE REGULATION OF MULTICELLULAR ORGANISMAL PROCESS | 16 | 1395 | 0.0002303 | 0.003133 |
| 343 | INTRACELLULAR STEROID HORMONE RECEPTOR SIGNALING PATHWAY | 4 | 71 | 0.0002377 | 0.003224 |
| 344 | CELL MATURATION | 5 | 131 | 0.0002408 | 0.003257 |
| 345 | SENSORY ORGAN DEVELOPMENT | 9 | 493 | 0.0002445 | 0.003297 |
| 346 | NEGATIVE REGULATION OF PRODUCTION OF MOLECULAR MEDIATOR OF IMMUNE RESPONSE | 3 | 29 | 0.0002499 | 0.003332 |
| 347 | REGULATION OF EXTRACELLULAR MATRIX ORGANIZATION | 3 | 29 | 0.0002499 | 0.003332 |
| 348 | REGULATION OF HEART MORPHOGENESIS | 3 | 29 | 0.0002499 | 0.003332 |
| 349 | POSITIVE REGULATION OF CELL CYCLE G1 S PHASE TRANSITION | 3 | 29 | 0.0002499 | 0.003332 |
| 350 | NEGATIVE REGULATION OF CELL DIFFERENTIATION | 10 | 609 | 0.0002556 | 0.003398 |
| 351 | REGULATION OF CELL GROWTH | 8 | 391 | 0.0002574 | 0.003412 |
| 352 | TRANSCRIPTION COUPLED NUCLEOTIDE EXCISION REPAIR | 4 | 73 | 0.0002645 | 0.003497 |
| 353 | RESPONSE TO ETHANOL | 5 | 136 | 0.0002865 | 0.003776 |
| 354 | REGULATION OF CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY | 6 | 213 | 0.0002954 | 0.003872 |
| 355 | CELL DEVELOPMENT | 16 | 1426 | 0.0002951 | 0.003872 |
| 356 | REGULATION OF PROTEIN TARGETING | 7 | 307 | 0.0003327 | 0.004349 |
| 357 | NEGATIVE REGULATION OF CYCLIN DEPENDENT PROTEIN KINASE ACTIVITY | 3 | 32 | 0.0003361 | 0.004369 |
| 358 | ENDOCARDIAL CUSHION DEVELOPMENT | 3 | 32 | 0.0003361 | 0.004369 |
| 359 | SOMITE DEVELOPMENT | 4 | 78 | 0.0003411 | 0.004421 |
| 360 | REGULATION OF GROWTH | 10 | 633 | 0.0003474 | 0.00449 |
| 361 | IN UTERO EMBRYONIC DEVELOPMENT | 7 | 311 | 0.0003597 | 0.004623 |
| 362 | NEGATIVE REGULATION OF CYTOSKELETON ORGANIZATION | 6 | 221 | 0.0003597 | 0.004623 |
| 363 | RAS PROTEIN SIGNAL TRANSDUCTION | 5 | 143 | 0.000361 | 0.004628 |
| 364 | REGULATION OF PROTEIN EXPORT FROM NUCLEUS | 3 | 33 | 0.0003686 | 0.004712 |
| 365 | EMBRYO DEVELOPMENT | 12 | 894 | 0.000385 | 0.004908 |
| 366 | RESPONSE TO ESTRADIOL | 5 | 146 | 0.0003972 | 0.005049 |
| 367 | HEART VALVE DEVELOPMENT | 3 | 34 | 0.0004031 | 0.00511 |
| 368 | CELLULAR CATABOLIC PROCESS | 15 | 1322 | 0.0004166 | 0.005267 |
| 369 | RESPONSE TO TRANSITION METAL NANOPARTICLE | 5 | 148 | 0.0004227 | 0.00533 |
| 370 | REGULATION OF GENE EXPRESSION EPIGENETIC | 6 | 229 | 0.0004344 | 0.005463 |
| 371 | RESPONSE TO MONOAMINE | 3 | 35 | 0.0004395 | 0.005512 |
| 372 | CHROMATIN REMODELING | 5 | 150 | 0.0004495 | 0.005622 |
| 373 | EYE DEVELOPMENT | 7 | 326 | 0.0004768 | 0.005948 |
| 374 | TRANSCRIPTION INITIATION FROM RNA POLYMERASE II PROMOTER | 5 | 153 | 0.000492 | 0.006121 |
| 375 | OVULATION CYCLE PROCESS | 4 | 88 | 0.00054 | 0.0067 |
| 376 | REGULATION OF MULTICELLULAR ORGANISMAL METABOLIC PROCESS | 3 | 38 | 0.0005613 | 0.006946 |
| 377 | SEGMENTATION | 4 | 89 | 0.0005636 | 0.006956 |
| 378 | RESPONSE TO TOXIC SUBSTANCE | 6 | 241 | 0.0005687 | 0.007001 |
| 379 | EMBRYO DEVELOPMENT ENDING IN BIRTH OR EGG HATCHING | 9 | 554 | 0.0005711 | 0.007012 |
| 380 | PROTEIN LOCALIZATION TO ORGANELLE | 9 | 556 | 0.0005861 | 0.007161 |
| 381 | REGULATION OF CELL MATRIX ADHESION | 4 | 90 | 0.0005879 | 0.007161 |
| 382 | MESONEPHROS DEVELOPMENT | 4 | 90 | 0.0005879 | 0.007161 |
| 383 | CELLULAR RESPONSE TO NUTRIENT | 3 | 39 | 0.0006062 | 0.007346 |
| 384 | SPLEEN DEVELOPMENT | 3 | 39 | 0.0006062 | 0.007346 |
| 385 | REGULATION OF CYTOKINE PRODUCTION | 9 | 563 | 0.0006409 | 0.007745 |
| 386 | POSITIVE REGULATION OF CELL MATRIX ADHESION | 3 | 40 | 0.0006534 | 0.007876 |
| 387 | CELLULAR MACROMOLECULE LOCALIZATION | 14 | 1234 | 0.0006645 | 0.00799 |
| 388 | COVALENT CHROMATIN MODIFICATION | 7 | 345 | 0.0006669 | 0.007997 |
| 389 | REGULATION OF DNA BIOSYNTHETIC PROCESS | 4 | 94 | 0.0006926 | 0.008285 |
| 390 | ANDROGEN RECEPTOR SIGNALING PATHWAY | 3 | 41 | 0.0007028 | 0.008385 |
| 391 | POSITIVE REGULATION OF CELL DIFFERENTIATION | 11 | 823 | 0.0007105 | 0.008455 |
| 392 | RESPONSE TO OXIDATIVE STRESS | 7 | 352 | 0.0007504 | 0.008907 |
| 393 | NEGATIVE REGULATION OF CELL GROWTH | 5 | 170 | 0.0007929 | 0.009388 |
| 394 | HEMATOPOIETIC PROGENITOR CELL DIFFERENTIATION | 4 | 98 | 0.00081 | 0.009539 |
| 395 | POSITIVE REGULATION OF INTRACELLULAR PROTEIN TRANSPORT | 6 | 258 | 0.0008119 | 0.009539 |
| 396 | BETA CATENIN TCF COMPLEX ASSEMBLY | 3 | 43 | 0.0008087 | 0.009539 |
| 397 | CYTOSKELETON ORGANIZATION | 11 | 838 | 0.000824 | 0.009658 |
| 398 | REGULATION OF CELL SIZE | 5 | 172 | 0.0008357 | 0.009768 |
| 399 | HEAD DEVELOPMENT | 10 | 709 | 0.0008376 | 0.009768 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | CYCLIN DEPENDENT PROTEIN SERINE THREONINE KINASE REGULATOR ACTIVITY | 9 | 28 | 1.892e-15 | 1.758e-12 |
| 2 | KINASE BINDING | 22 | 606 | 5.693e-15 | 2.644e-12 |
| 3 | PROTEIN KINASE ACTIVITY | 22 | 640 | 1.738e-14 | 5.383e-12 |
| 4 | ENZYME BINDING | 33 | 1737 | 3.249e-14 | 7.547e-12 |
| 5 | TRANSCRIPTION FACTOR BINDING | 20 | 524 | 4.697e-14 | 8.727e-12 |
| 6 | PROTEIN SERINE THREONINE KINASE ACTIVITY | 18 | 445 | 3.896e-13 | 6.032e-11 |
| 7 | KINASE ACTIVITY | 23 | 842 | 4.936e-13 | 6.551e-11 |
| 8 | TRANSFERASE ACTIVITY TRANSFERRING PHOSPHORUS CONTAINING GROUPS | 23 | 992 | 1.411e-11 | 1.638e-09 |
| 9 | CYCLIN DEPENDENT PROTEIN KINASE ACTIVITY | 7 | 34 | 9.543e-11 | 9.851e-09 |
| 10 | PROTEIN COMPLEX BINDING | 20 | 935 | 1.592e-09 | 1.479e-07 |
| 11 | KINASE REGULATOR ACTIVITY | 10 | 186 | 6.498e-09 | 5.488e-07 |
| 12 | MACROMOLECULAR COMPLEX BINDING | 23 | 1399 | 1.155e-08 | 8.94e-07 |
| 13 | ADENYL NUCLEOTIDE BINDING | 23 | 1514 | 5.006e-08 | 3.577e-06 |
| 14 | CYCLIN DEPENDENT PROTEIN SERINE THREONINE KINASE INHIBITOR ACTIVITY | 4 | 12 | 1.465e-07 | 9.723e-06 |
| 15 | PROTEIN SERINE THREONINE KINASE INHIBITOR ACTIVITY | 5 | 30 | 1.613e-07 | 9.992e-06 |
| 16 | RNA POLYMERASE II TRANSCRIPTION FACTOR BINDING | 7 | 104 | 2.967e-07 | 1.723e-05 |
| 17 | CYCLIN BINDING | 4 | 19 | 1.122e-06 | 6.129e-05 |
| 18 | STEROID HORMONE RECEPTOR BINDING | 6 | 81 | 1.239e-06 | 6.396e-05 |
| 19 | RIBONUCLEOTIDE BINDING | 23 | 1860 | 1.943e-06 | 9.502e-05 |
| 20 | DNA HELICASE ACTIVITY | 5 | 53 | 3.009e-06 | 0.0001398 |
| 21 | CORE PROMOTER BINDING | 7 | 152 | 3.833e-06 | 0.0001695 |
| 22 | ACTIVATING TRANSCRIPTION FACTOR BINDING | 5 | 57 | 4.332e-06 | 0.0001829 |
| 23 | NF KAPPAB BINDING | 4 | 30 | 7.653e-06 | 0.0003091 |
| 24 | NUCLEIC ACID BINDING TRANSCRIPTION FACTOR ACTIVITY | 17 | 1199 | 9.818e-06 | 0.0003648 |
| 25 | P53 BINDING | 5 | 67 | 9.666e-06 | 0.0003648 |
| 26 | ENZYME REGULATOR ACTIVITY | 15 | 959 | 1.127e-05 | 0.0004027 |
| 27 | SMAD BINDING | 5 | 72 | 1.377e-05 | 0.0004739 |
| 28 | UBIQUITIN LIKE PROTEIN LIGASE BINDING | 8 | 264 | 1.659e-05 | 0.0005137 |
| 29 | RNA POLYMERASE II ACTIVATING TRANSCRIPTION FACTOR BINDING | 4 | 36 | 1.613e-05 | 0.0005137 |
| 30 | CHROMATIN BINDING | 10 | 435 | 1.56e-05 | 0.0005137 |
| 31 | ANDROGEN RECEPTOR BINDING | 4 | 39 | 2.231e-05 | 0.0006477 |
| 32 | DNA DEPENDENT ATPASE ACTIVITY | 5 | 79 | 2.167e-05 | 0.0006477 |
| 33 | LIGASE REGULATOR ACTIVITY | 3 | 14 | 2.607e-05 | 0.0007339 |
| 34 | PEROXISOME PROLIFERATOR ACTIVATED RECEPTOR BINDING | 3 | 15 | 3.249e-05 | 0.0008877 |
| 35 | KINASE INHIBITOR ACTIVITY | 5 | 89 | 3.861e-05 | 0.00102 |
| 36 | REGULATORY REGION NUCLEIC ACID BINDING | 13 | 818 | 3.951e-05 | 0.00102 |
| 37 | MOLECULAR FUNCTION REGULATOR | 17 | 1353 | 4.622e-05 | 0.001161 |
| 38 | TRANSFORMING GROWTH FACTOR BETA RECEPTOR BINDING | 4 | 50 | 6.028e-05 | 0.001474 |
| 39 | HISTONE KINASE ACTIVITY | 3 | 19 | 6.834e-05 | 0.001628 |
| 40 | RNA POLYMERASE II TRANSCRIPTION FACTOR ACTIVITY SEQUENCE SPECIFIC DNA BINDING | 11 | 629 | 7.049e-05 | 0.001637 |
| 41 | HORMONE RECEPTOR BINDING | 6 | 168 | 8.119e-05 | 0.00184 |
| 42 | R SMAD BINDING | 3 | 23 | 0.0001234 | 0.002729 |
| 43 | PROTEIN HETERODIMERIZATION ACTIVITY | 9 | 468 | 0.0001661 | 0.003589 |
| 44 | CORE PROMOTER PROXIMAL REGION DNA BINDING | 8 | 371 | 0.0001804 | 0.003809 |
| 45 | TRANSCRIPTION FACTOR ACTIVITY PROTEIN BINDING | 10 | 588 | 0.0001929 | 0.003983 |
| 46 | BHLH TRANSCRIPTION FACTOR BINDING | 3 | 28 | 0.0002248 | 0.004539 |
| 47 | PROTEIN DOMAIN SPECIFIC BINDING | 10 | 624 | 0.0003102 | 0.006132 |
| 48 | TRANSCRIPTIONAL ACTIVATOR ACTIVITY RNA POLYMERASE II TRANSCRIPTION REGULATORY REGION SEQUENCE SPECIFIC BINDING | 7 | 315 | 0.0003884 | 0.007363 |
| 49 | DOUBLE STRANDED DNA BINDING | 11 | 764 | 0.0003821 | 0.007363 |
| 50 | ATP DEPENDENT DNA HELICASE ACTIVITY | 3 | 34 | 0.0004031 | 0.007489 |
| 51 | RECEPTOR BINDING | 16 | 1476 | 0.0004333 | 0.007892 |
| 52 | BETA CATENIN BINDING | 4 | 84 | 0.0004526 | 0.008086 |
| 53 | HELICASE ACTIVITY | 5 | 153 | 0.000492 | 0.008624 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | CHROMOSOME | 33 | 880 | 4.328e-23 | 2.528e-20 |
| 2 | CHROMOSOMAL REGION | 21 | 330 | 3.265e-19 | 9.534e-17 |
| 3 | TRANSCRIPTION FACTOR COMPLEX | 20 | 298 | 9.022e-19 | 1.756e-16 |
| 4 | NUCLEAR CHROMOSOME | 23 | 523 | 1.905e-17 | 2.781e-15 |
| 5 | CYCLIN DEPENDENT PROTEIN KINASE HOLOENZYME COMPLEX | 10 | 31 | 4.588e-17 | 5.359e-15 |
| 6 | CHROMATIN | 20 | 441 | 1.809e-15 | 1.761e-13 |
| 7 | TRANSFERASE COMPLEX | 23 | 703 | 1.124e-14 | 9.373e-13 |
| 8 | PROTEIN KINASE COMPLEX | 11 | 90 | 1.326e-13 | 9.68e-12 |
| 9 | MICROTUBULE CYTOSKELETON | 25 | 1068 | 1.176e-12 | 7.629e-11 |
| 10 | CATALYTIC COMPLEX | 23 | 1038 | 3.508e-11 | 2.049e-09 |
| 11 | CHROMOSOME CENTROMERIC REGION | 11 | 174 | 1.922e-10 | 1.021e-08 |
| 12 | SPINDLE | 13 | 289 | 2.669e-10 | 1.299e-08 |
| 13 | TRANSFERASE COMPLEX TRANSFERRING PHOSPHORUS CONTAINING GROUPS | 12 | 237 | 3.559e-10 | 1.485e-08 |
| 14 | ANAPHASE PROMOTING COMPLEX | 6 | 22 | 3.48e-10 | 1.485e-08 |
| 15 | MCM COMPLEX | 5 | 11 | 5.573e-10 | 2.17e-08 |
| 16 | CONDENSED CHROMOSOME | 11 | 195 | 6.493e-10 | 2.37e-08 |
| 17 | CHROMOSOME TELOMERIC REGION | 10 | 162 | 1.708e-09 | 5.866e-08 |
| 18 | CENTROSOME | 15 | 487 | 1.908e-09 | 6.19e-08 |
| 19 | CONDENSED NUCLEAR CHROMOSOME | 8 | 85 | 2.805e-09 | 8.622e-08 |
| 20 | MICROTUBULE ORGANIZING CENTER | 16 | 623 | 6.953e-09 | 2.03e-07 |
| 21 | CYTOSKELETON | 27 | 1967 | 2.027e-08 | 5.637e-07 |
| 22 | NUCLEAR UBIQUITIN LIGASE COMPLEX | 6 | 42 | 2.286e-08 | 6.069e-07 |
| 23 | SPINDLE POLE | 8 | 126 | 6.36e-08 | 1.615e-06 |
| 24 | NUCLEAR CHROMOSOME TELOMERIC REGION | 8 | 132 | 9.135e-08 | 2.134e-06 |
| 25 | CYTOSKELETAL PART | 22 | 1436 | 9.12e-08 | 2.134e-06 |
| 26 | CONDENSED CHROMOSOME OUTER KINETOCHORE | 4 | 12 | 1.465e-07 | 3.291e-06 |
| 27 | CULLIN RING UBIQUITIN LIGASE COMPLEX | 8 | 150 | 2.452e-07 | 5.303e-06 |
| 28 | CONDENSED CHROMOSOME CENTROMERIC REGION | 7 | 102 | 2.597e-07 | 5.416e-06 |
| 29 | KINETOCHORE | 7 | 120 | 7.866e-07 | 1.584e-05 |
| 30 | CONDENSED NUCLEAR CHROMOSOME CENTROMERIC REGION | 4 | 18 | 8.884e-07 | 1.729e-05 |
| 31 | RNA POLYMERASE II TRANSCRIPTION FACTOR COMPLEX | 6 | 101 | 4.523e-06 | 8.521e-05 |
| 32 | COHESIN COMPLEX | 3 | 11 | 1.193e-05 | 0.0002177 |
| 33 | NUCLEOLUS | 14 | 848 | 1.258e-05 | 0.0002227 |
| 34 | UBIQUITIN LIGASE COMPLEX | 8 | 262 | 1.57e-05 | 0.0002697 |
| 35 | NUCLEAR TRANSCRIPTION FACTOR COMPLEX | 6 | 127 | 1.69e-05 | 0.0002819 |
| 36 | CARBOXY TERMINAL DOMAIN PROTEIN KINASE COMPLEX | 3 | 22 | 0.0001076 | 0.001746 |
| 37 | SPINDLE MIDZONE | 3 | 27 | 0.0002013 | 0.003177 |
| 38 | NUCLEAR CHROMATIN | 7 | 291 | 0.0002406 | 0.003698 |
Over-represented Pathway
| Num | Pathway | Pathview | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|---|
| 1 | hsa04110_Cell_cycle | 85 | 128 | 1.973e-203 | 3.551e-201 | |
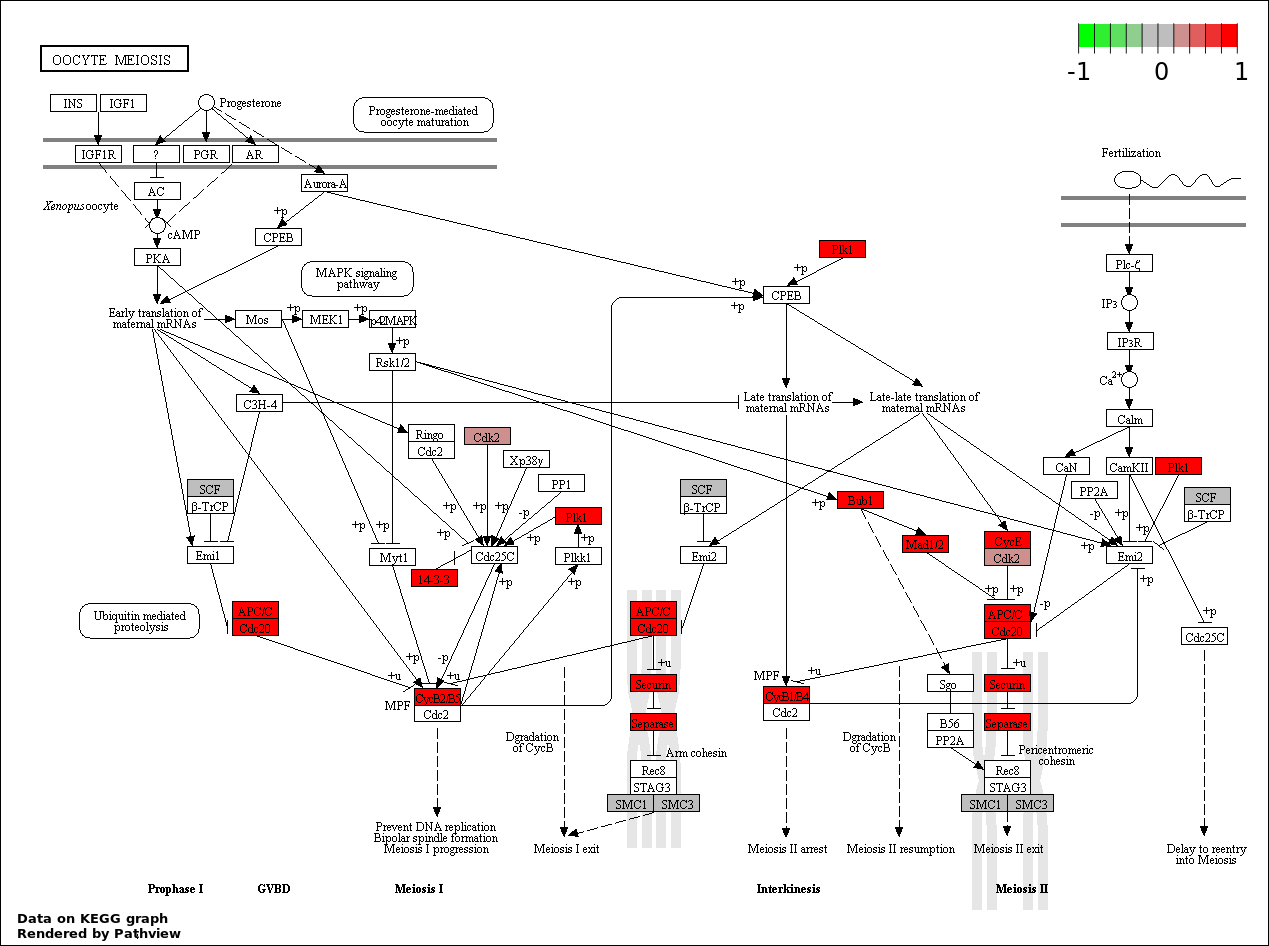
| 2 | hsa04114_Oocyte_meiosis | 22 | 114 | 4.72e-31 | 4.248e-29 | |
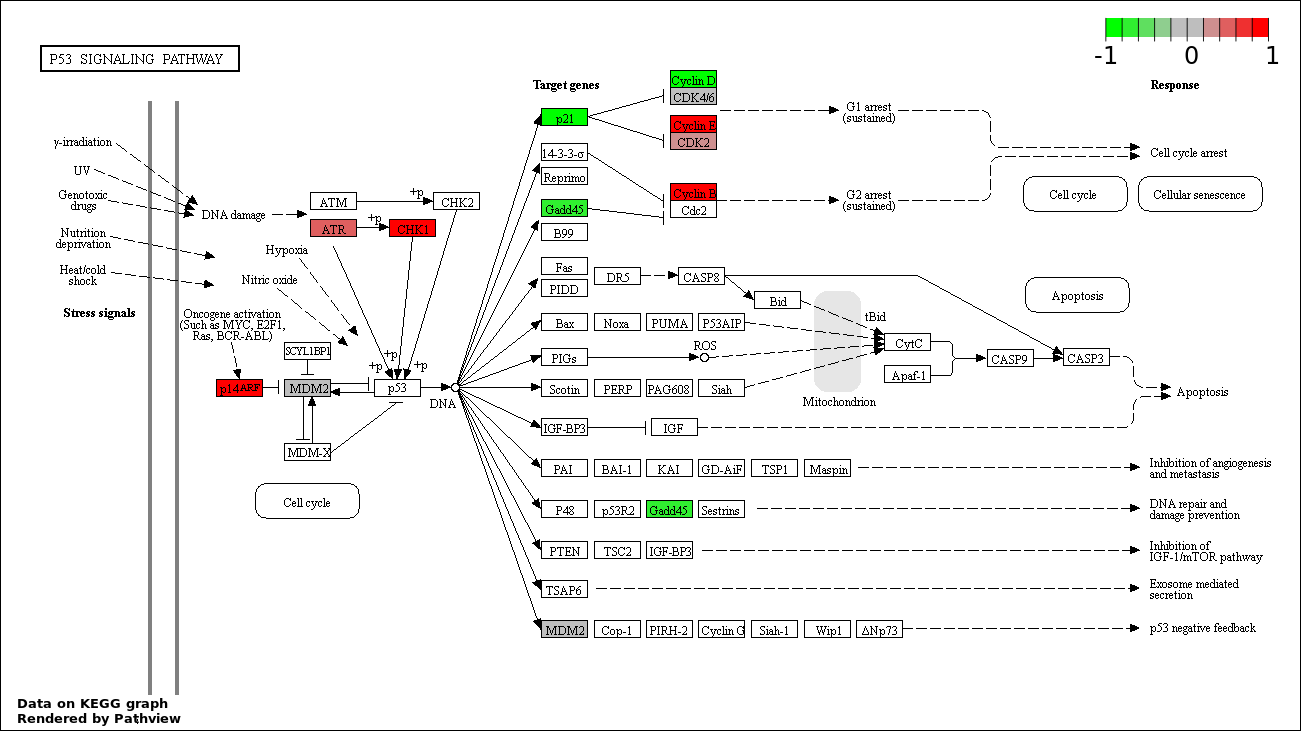
| 3 | hsa04115_p53_signaling_pathway | 18 | 69 | 4.349e-28 | 2.61e-26 | |
| 4 | hsa04350_TGF.beta_signaling_pathway | 15 | 85 | 1.03e-20 | 4.637e-19 | |
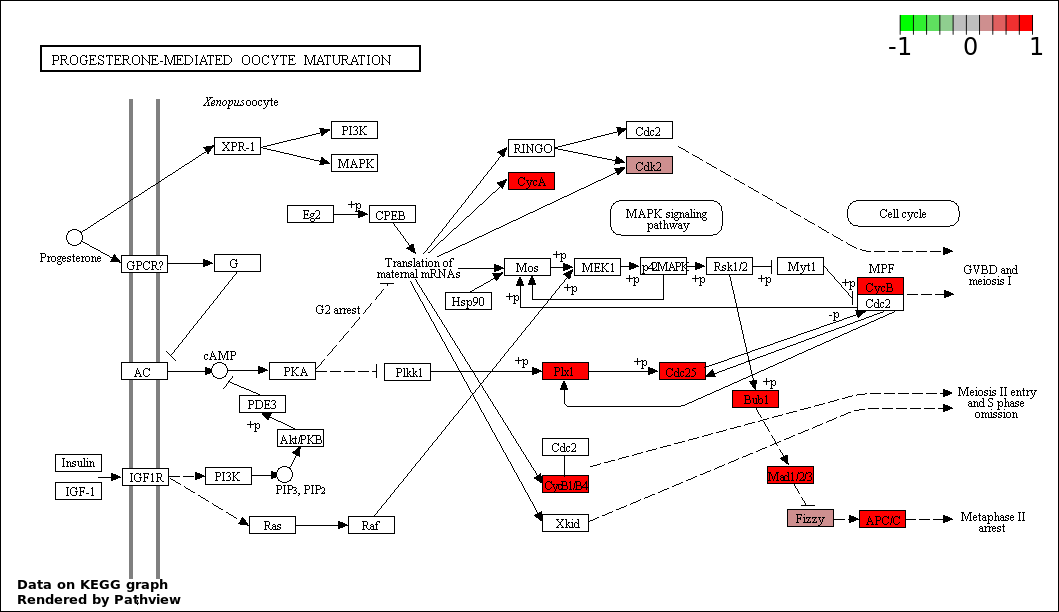
| 5 | hsa04914_Progesterone.mediated_oocyte_maturation | 14 | 87 | 8.617e-19 | 3.102e-17 | |
| 6 | hsa04390_Hippo_signaling_pathway | 15 | 154 | 1.121e-16 | 3.363e-15 | |
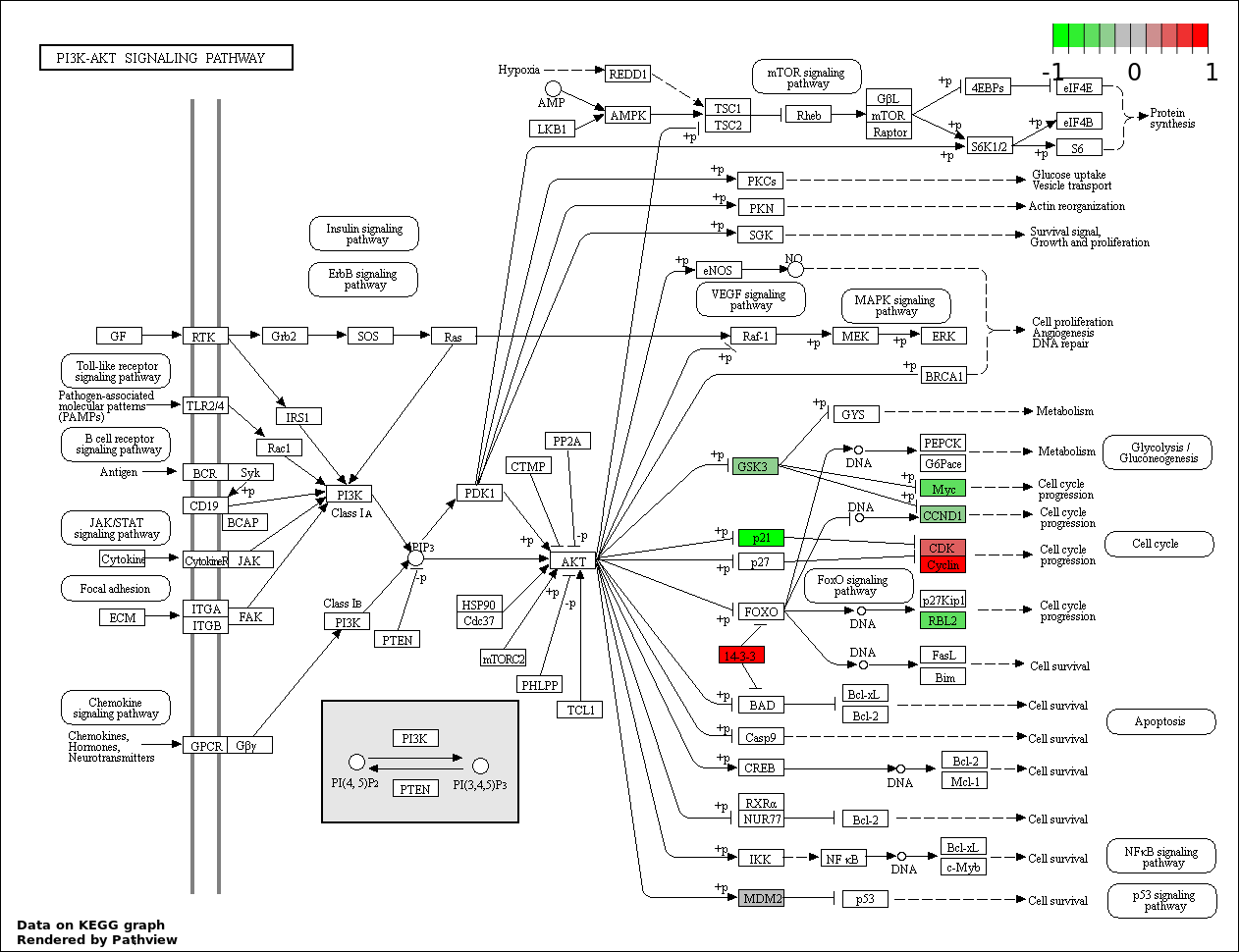
| 7 | hsa04151_PI3K_AKT_signaling_pathway | 18 | 351 | 6.702e-15 | 1.723e-13 | |
| 8 | hsa04310_Wnt_signaling_pathway | 11 | 151 | 4.156e-11 | 9.352e-10 | |
| 9 | hsa03030_DNA_replication | 6 | 36 | 8.663e-09 | 1.733e-07 | |
| 10 | hsa04120_Ubiquitin_mediated_proteolysis | 8 | 139 | 1.363e-07 | 2.454e-06 | |
| 11 | hsa04722_Neurotrophin_signaling_pathway | 7 | 127 | 1.154e-06 | 1.888e-05 | |
| 12 | hsa04520_Adherens_junction | 5 | 73 | 1.474e-05 | 0.000221 | |
| 13 | hsa04630_Jak.STAT_signaling_pathway | 6 | 155 | 5.193e-05 | 0.000719 | |
| 14 | hsa04010_MAPK_signaling_pathway | 7 | 268 | 0.0001453 | 0.001868 | |
| 15 | hsa04012_ErbB_signaling_pathway | 4 | 87 | 0.0005171 | 0.006205 | |
| 16 | hsa03420_Nucleotide_excision_repair | 3 | 45 | 0.0009242 | 0.0104 | |
| 17 | hsa04330_Notch_signaling_pathway | 3 | 47 | 0.00105 | 0.01111 | |
| 18 | hsa04144_Endocytosis | 5 | 203 | 0.001743 | 0.01743 | |
| 19 | hsa04916_Melanogenesis | 3 | 101 | 0.009153 | 0.08671 | |
| 20 | hsa04510_Focal_adhesion | 4 | 200 | 0.01047 | 0.09421 | |
| 21 | hsa04720_Long.term_potentiation | 2 | 70 | 0.03578 | 0.3067 | |
| 22 | hsa04660_T_cell_receptor_signaling_pathway | 2 | 108 | 0.07727 | 0.6047 | |
| 23 | hsa04360_Axon_guidance | 2 | 130 | 0.1057 | 0.7321 |


















lncRNA-mediated sponge
| Num | lncRNA | miRNAs | miRNAs count | Gene | Sponge regulatory network | lncRNA log2FC | lncRNA pvalue | Gene log2FC | Gene pvalue | lncRNA-gene Pearson correlation |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | RP1-228H13.5 | hsa-let-7b-5p;hsa-miR-125a-5p;hsa-miR-125b-5p;hsa-miR-140-3p;hsa-miR-145-5p;hsa-miR-195-5p;hsa-miR-221-3p;hsa-miR-30a-5p;hsa-miR-30c-5p;hsa-miR-30d-5p;hsa-miR-320a | 11 | E2F3 | Sponge network | 1.701 | 0 | 1.313 | 0 | 0.538 |
| 2 | RP11-389C8.2 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-550a-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-7-1-3p;hsa-miR-93-5p | 24 | CCND2 | Sponge network | -2.039 | 0 | -1.641 | 0 | 0.52 |
| 3 | AF131215.2 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-191-5p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-301a-3p;hsa-miR-324-3p;hsa-miR-550a-5p;hsa-miR-589-3p | 14 | CCND2 | Sponge network | -2.09 | 0 | -1.641 | 0 | 0.505 |
| 4 | LINC00337 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7f-5p;hsa-let-7g-5p;hsa-miR-125a-5p;hsa-miR-26a-5p;hsa-miR-30c-2-3p;hsa-miR-320a;hsa-miR-338-3p | 10 | E2F2 | Sponge network | 2.531 | 1.0E-5 | 2.225 | 0 | 0.495 |
| 5 | RP11-750H9.5 | hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-2355-3p;hsa-miR-301a-3p;hsa-miR-550a-5p;hsa-miR-877-5p | 10 | CCND2 | Sponge network | -1.959 | 0 | -1.641 | 0 | 0.493 |
| 6 | PCED1B-AS1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-877-5p;hsa-miR-93-5p;hsa-miR-96-5p | 18 | CCND2 | Sponge network | -0.672 | 0.02084 | -1.641 | 0 | 0.488 |
| 7 | RP11-399O19.9 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-7-1-3p;hsa-miR-93-5p | 18 | CCND2 | Sponge network | -0.873 | 0.00072 | -1.641 | 0 | 0.469 |
| 8 | AF131215.9 | hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-450b-5p;hsa-miR-503-5p;hsa-miR-590-3p | 13 | CCND2 | Sponge network | -1.808 | 0 | -1.641 | 0 | 0.46 |
| 9 | BZRAP1-AS1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-550a-5p;hsa-miR-877-5p | 14 | CCND2 | Sponge network | -0.785 | 0.00723 | -1.641 | 0 | 0.456 |
| 10 | LINC00996 | hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-320b;hsa-miR-550a-5p | 10 | CCND2 | Sponge network | -1.372 | 0.00025 | -1.641 | 0 | 0.454 |
| 11 | RP11-532F6.3 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-301a-3p | 13 | CCND2 | Sponge network | -2.028 | 0 | -1.641 | 0 | 0.445 |
| 12 | DENND5B-AS1 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-let-7g-5p;hsa-miR-125a-5p;hsa-miR-125b-5p;hsa-miR-26a-5p;hsa-miR-30c-2-3p;hsa-miR-320a | 10 | E2F2 | Sponge network | 1.989 | 0 | 2.225 | 0 | 0.436 |
| 13 | LINC00702 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-324-3p;hsa-miR-450b-5p;hsa-miR-501-5p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-660-5p;hsa-miR-7-1-3p;hsa-miR-877-5p;hsa-miR-93-5p;hsa-miR-96-5p | 34 | CCND2 | Sponge network | -2.856 | 0 | -1.641 | 0 | 0.433 |
| 14 | LINC00702 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-454-3p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p | 16 | TGFB2 | Sponge network | -2.856 | 0 | -1.547 | 1.0E-5 | 0.413 |
| 15 | TBX5-AS1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-324-3p;hsa-miR-450b-5p;hsa-miR-501-5p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-877-5p | 26 | CCND2 | Sponge network | -2.108 | 0 | -1.641 | 0 | 0.404 |
| 16 | AC011899.9 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-590-3p;hsa-miR-7-1-3p | 19 | CCND2 | Sponge network | -2.611 | 0 | -1.641 | 0 | 0.404 |
| 17 | RP11-253E3.3 | hsa-miR-141-3p;hsa-miR-148a-3p;hsa-miR-15a-5p;hsa-miR-182-5p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-21-5p;hsa-miR-27b-5p;hsa-miR-29b-3p;hsa-miR-30e-5p;hsa-miR-660-5p | 11 | CDK6 | Sponge network | -2.404 | 0 | -0.635 | 0.01747 | 0.403 |
| 18 | LINC00968 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-324-3p;hsa-miR-450b-5p;hsa-miR-503-5p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-93-5p;hsa-miR-96-5p | 27 | CCND2 | Sponge network | -4.19 | 0 | -1.641 | 0 | 0.396 |
| 19 | RP11-720L2.4 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-93-5p | 12 | CCND2 | Sponge network | -2.305 | 0 | -1.641 | 0 | 0.394 |
| 20 | RNF144A-AS1 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7f-5p;hsa-let-7g-5p;hsa-miR-125a-5p;hsa-miR-125b-5p;hsa-miR-26a-5p;hsa-miR-3065-5p;hsa-miR-30c-2-3p;hsa-miR-320a | 11 | E2F2 | Sponge network | 1.592 | 0.00021 | 2.225 | 0 | 0.38 |
| 21 | MAGI2-AS3 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-429;hsa-miR-501-5p;hsa-miR-503-5p;hsa-miR-590-3p;hsa-miR-660-5p;hsa-miR-7-1-3p;hsa-miR-877-5p;hsa-miR-93-5p | 27 | CCND2 | Sponge network | -1.892 | 0 | -1.641 | 0 | 0.378 |
| 22 | RP11-1024P17.1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-429;hsa-miR-589-3p;hsa-miR-590-3p | 21 | CCND2 | Sponge network | -2.062 | 0 | -1.641 | 0 | 0.374 |
| 23 | RP5-1120P11.1 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-let-7e-5p;hsa-let-7f-5p;hsa-let-7g-5p;hsa-miR-125a-5p;hsa-miR-26a-5p;hsa-miR-338-3p | 10 | E2F2 | Sponge network | 1.924 | 1.0E-5 | 2.225 | 0 | 0.369 |
| 24 | RP11-1008C21.1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-550a-5p;hsa-miR-590-3p | 17 | CCND2 | Sponge network | -1.826 | 3.0E-5 | -1.641 | 0 | 0.36 |
| 25 | RP11-284N8.3 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-183-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-324-3p;hsa-miR-331-5p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-93-5p;hsa-miR-96-5p | 20 | CCND2 | Sponge network | -0.761 | 0.05061 | -1.641 | 0 | 0.36 |
| 26 | RP11-354E11.2 |
hsa-miR-107;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-590-3p | 10 | TGFB2 | Sponge network | -2.138 | 0 | -1.547 | 1.0E-5 | 0.356 |
| 27 | RP11-1024P17.1 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-335-3p;hsa-miR-589-3p;hsa-miR-590-3p | 13 | TGFB2 | Sponge network | -2.062 | 0 | -1.547 | 1.0E-5 | 0.354 |
| 28 | CTD-2013N24.2 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-501-5p;hsa-miR-590-3p;hsa-miR-660-5p;hsa-miR-7-1-3p;hsa-miR-93-5p | 24 | CCND2 | Sponge network | -1.745 | 0 | -1.641 | 0 | 0.354 |
| 29 | AC011526.1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-450b-5p;hsa-miR-503-5p;hsa-miR-93-5p;hsa-miR-96-5p | 10 | CCND2 | Sponge network | -2.783 | 0 | -1.641 | 0 | 0.352 |
| 30 | CTD-2003C8.2 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-550a-5p | 11 | CCND2 | Sponge network | -3.403 | 0 | -1.641 | 0 | 0.345 |
| 31 | AC109642.1 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-590-3p | 11 | TGFB2 | Sponge network | -2.791 | 0 | -1.547 | 1.0E-5 | 0.343 |
| 32 | MAGI2-AS3 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-454-3p;hsa-miR-590-3p | 14 | TGFB2 | Sponge network | -1.892 | 0 | -1.547 | 1.0E-5 | 0.342 |
| 33 | RP11-23P13.6 | hsa-miR-106b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-20a-5p;hsa-miR-324-3p;hsa-miR-331-5p;hsa-miR-590-3p;hsa-miR-877-5p | 11 | CCND2 | Sponge network | -0.705 | 0.00072 | -1.641 | 0 | 0.34 |
| 34 | AC109642.1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-590-3p;hsa-miR-93-5p;hsa-miR-96-5p | 24 | CCND2 | Sponge network | -2.791 | 0 | -1.641 | 0 | 0.339 |
| 35 | RP11-401P9.4 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-19a-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-550a-5p;hsa-miR-590-5p;hsa-miR-93-5p | 19 | CCND2 | Sponge network | -3.04 | 0 | -1.641 | 0 | 0.337 |
| 36 | GAS6-AS2 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-550a-5p;hsa-miR-590-5p;hsa-miR-7-1-3p | 23 | CCND2 | Sponge network | -1.761 | 0 | -1.641 | 0 | 0.337 |
| 37 | RP11-354E11.2 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-590-3p;hsa-miR-96-5p | 17 | CCND2 | Sponge network | -2.138 | 0 | -1.641 | 0 | 0.335 |
| 38 | CTD-2008P7.8 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-let-7f-5p;hsa-let-7g-5p;hsa-miR-125a-5p;hsa-miR-125b-5p;hsa-miR-30c-2-3p;hsa-miR-338-3p | 10 | E2F2 | Sponge network | 5.852 | 0 | 2.225 | 0 | 0.335 |
| 39 | CTD-2013N24.2 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-454-3p;hsa-miR-590-3p | 13 | TGFB2 | Sponge network | -1.745 | 0 | -1.547 | 1.0E-5 | 0.334 |
| 40 | LINC00472 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-301a-3p;hsa-miR-450b-5p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-7-1-3p;hsa-miR-93-5p | 18 | CCND2 | Sponge network | -2.952 | 0 | -1.641 | 0 | 0.327 |
| 41 | AC144831.1 | hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-20a-5p;hsa-miR-28-5p;hsa-miR-324-3p;hsa-miR-550a-5p;hsa-miR-7-1-3p;hsa-miR-877-5p;hsa-miR-93-5p | 14 | CCND2 | Sponge network | -2.063 | 0 | -1.641 | 0 | 0.327 |
| 42 | RP11-166D19.1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-324-3p;hsa-miR-429;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-877-5p;hsa-miR-96-5p | 22 | CCND2 | Sponge network | -0.582 | 0.05253 | -1.641 | 0 | 0.322 |
| 43 | LINC00092 |
hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-503-5p;hsa-miR-589-3p;hsa-miR-590-3p | 14 | CCND2 | Sponge network | -2.383 | 0 | -1.641 | 0 | 0.321 |
| 44 | TBX5-AS1 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-335-3p;hsa-miR-590-5p | 12 | TGFB2 | Sponge network | -2.108 | 0 | -1.547 | 1.0E-5 | 0.32 |
| 45 | RP5-1042I8.7 | hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-20a-5p;hsa-miR-320b;hsa-miR-331-5p;hsa-miR-590-3p;hsa-miR-93-5p;hsa-miR-96-5p | 13 | CCND2 | Sponge network | -0.733 | 0.00018 | -1.641 | 0 | 0.316 |
| 46 | AC079630.4 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-320b;hsa-miR-331-5p;hsa-miR-550a-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-7-1-3p;hsa-miR-877-5p;hsa-miR-93-5p | 19 | CCND2 | Sponge network | -3.758 | 0 | -1.641 | 0 | 0.314 |
| 47 | LINC00968 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-590-3p;hsa-miR-590-5p | 12 | TGFB2 | Sponge network | -4.19 | 0 | -1.547 | 1.0E-5 | 0.308 |
| 48 | DIO3OS | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-301a-3p;hsa-miR-324-3p;hsa-miR-331-5p;hsa-miR-550a-5p;hsa-miR-877-5p | 14 | CCND2 | Sponge network | -1.936 | 0.00085 | -1.641 | 0 | 0.307 |
| 49 | RP11-283G6.4 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-450b-5p;hsa-miR-93-5p;hsa-miR-96-5p | 12 | CCND2 | Sponge network | -3.669 | 1.0E-5 | -1.641 | 0 | 0.305 |
| 50 | CTC-366B18.4 | hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-320b;hsa-miR-590-3p;hsa-miR-93-5p | 10 | CCND2 | Sponge network | -0.652 | 0.01265 | -1.641 | 0 | 0.303 |
| 51 | FENDRR |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-331-5p;hsa-miR-450b-5p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-590-3p;hsa-miR-7-1-3p;hsa-miR-93-5p;hsa-miR-96-5p | 22 | CCND2 | Sponge network | -4.222 | 0 | -1.641 | 0 | 0.301 |
| 52 | RP11-378A13.1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-590-3p;hsa-miR-877-5p;hsa-miR-93-5p | 20 | CCND2 | Sponge network | -1.713 | 0 | -1.641 | 0 | 0.301 |
| 53 | BAIAP2-AS1 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-142-3p;hsa-miR-17-5p;hsa-miR-193a-3p;hsa-miR-193b-3p;hsa-miR-20a-5p;hsa-miR-20b-5p;hsa-miR-365a-3p;hsa-miR-590-3p;hsa-miR-93-5p;hsa-miR-942-5p | 12 | CCND1 | Sponge network | -0.182 | 0.51705 | -0.296 | 0.2554 | 0.3 |
| 54 | RP4-647J21.1 | hsa-miR-106b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-183-5p;hsa-miR-200a-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-550a-5p;hsa-miR-93-5p | 10 | CCND2 | Sponge network | -0.153 | 0.73575 | -1.641 | 0 | 0.299 |
| 55 | LINC00922 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-324-3p;hsa-miR-501-5p;hsa-miR-7-1-3p;hsa-miR-877-5p | 14 | CCND2 | Sponge network | -0.842 | 0.11239 | -1.641 | 0 | 0.298 |
| 56 | RP11-476D10.1 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-550a-5p;hsa-miR-590-3p;hsa-miR-7-1-3p;hsa-miR-96-5p | 19 | CCND2 | Sponge network | -4.519 | 0 | -1.641 | 0 | 0.296 |
| 57 | RP11-416I2.1 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-15b-5p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-19a-3p;hsa-miR-19b-1-5p;hsa-miR-20a-5p;hsa-miR-20b-5p;hsa-miR-769-3p;hsa-miR-92a-3p;hsa-miR-93-5p;hsa-miR-942-5p | 13 | CCND1 | Sponge network | 3.177 | 1.0E-5 | -0.296 | 0.2554 | 0.294 |
| 58 | RP11-456K23.1 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-450b-5p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-93-5p | 23 | CCND2 | Sponge network | -1.488 | 0 | -1.641 | 0 | 0.292 |
| 59 | KTN1-AS1 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-miR-125a-5p;hsa-miR-125b-5p;hsa-miR-26a-5p;hsa-miR-3065-5p;hsa-miR-30c-2-3p;hsa-miR-320a;hsa-miR-338-3p | 10 | E2F2 | Sponge network | 1.013 | 0.00064 | 2.225 | 0 | 0.292 |
| 60 | LINC00961 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-589-3p;hsa-miR-96-5p | 16 | CCND2 | Sponge network | -2.724 | 0 | -1.641 | 0 | 0.292 |
| 61 | RP11-400K9.4 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-301a-3p;hsa-miR-93-5p | 12 | CCND2 | Sponge network | -1.193 | 0.00359 | -1.641 | 0 | 0.29 |
| 62 | AC004947.2 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-301a-3p;hsa-miR-450b-5p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-877-5p;hsa-miR-93-5p | 17 | CCND2 | Sponge network | -3.94 | 0 | -1.641 | 0 | 0.289 |
| 63 | AC006129.1 | hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-877-5p | 11 | CCND2 | Sponge network | -1.587 | 0.00086 | -1.641 | 0 | 0.287 |
| 64 | SH3RF3-AS1 | hsa-miR-15a-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-21-3p;hsa-miR-503-5p;hsa-miR-590-5p;hsa-miR-7-1-3p | 12 | CCND2 | Sponge network | -1.583 | 0 | -1.641 | 0 | 0.286 |
| 65 | RP3-508I15.9 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-let-7e-5p;hsa-let-7f-5p;hsa-let-7g-5p;hsa-miR-125b-5p;hsa-miR-26a-5p;hsa-miR-30c-2-3p | 10 | E2F2 | Sponge network | 1.462 | 0 | 2.225 | 0 | 0.284 |
| 66 | CTD-2357A8.3 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-let-7g-5p;hsa-miR-125a-5p;hsa-miR-125b-5p;hsa-miR-26a-5p;hsa-miR-30c-2-3p;hsa-miR-320a;hsa-miR-338-3p | 11 | E2F2 | Sponge network | 2.725 | 0 | 2.225 | 0 | 0.273 |
| 67 | AC010226.4 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-93-5p | 10 | CCND2 | Sponge network | -1.081 | 2.0E-5 | -1.641 | 0 | 0.272 |
| 68 | RP11-389C8.2 |
hsa-miR-107;hsa-miR-141-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-589-3p;hsa-miR-590-3p | 10 | TGFB2 | Sponge network | -2.039 | 0 | -1.547 | 1.0E-5 | 0.272 |
| 69 | MIR497HG |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-301a-3p;hsa-miR-324-3p;hsa-miR-503-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-877-5p;hsa-miR-93-5p;hsa-miR-96-5p | 27 | CCND2 | Sponge network | -2.142 | 0 | -1.641 | 0 | 0.271 |
| 70 | DNM3OS |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-183-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-191-5p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-501-5p;hsa-miR-550a-5p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-93-5p | 16 | CCND2 | Sponge network | 0.053 | 0.85755 | -1.641 | 0 | 0.27 |
| 71 | C1orf132 | hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-151a-3p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-21-3p;hsa-miR-2355-3p;hsa-miR-28-5p;hsa-miR-320b;hsa-miR-450b-5p;hsa-miR-503-5p;hsa-miR-550a-5p;hsa-miR-590-3p;hsa-miR-877-5p;hsa-miR-93-5p | 15 | CCND2 | Sponge network | -0.86 | 0.02429 | -1.641 | 0 | 0.269 |
| 72 | RP11-20J15.3 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-130b-5p;hsa-miR-17-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-550a-5p;hsa-miR-877-5p | 11 | CCND2 | Sponge network | -1.709 | 0.0268 | -1.641 | 0 | 0.264 |
| 73 | AC022182.3 | hsa-miR-106b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-2355-3p;hsa-miR-877-5p;hsa-miR-93-5p | 10 | CCND2 | Sponge network | -0.559 | 0.20451 | -1.641 | 0 | 0.263 |
| 74 | RP11-536K7.3 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-141-3p;hsa-miR-151a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-20a-5p;hsa-miR-877-5p;hsa-miR-93-5p | 13 | CCND2 | Sponge network | -1.239 | 5.0E-5 | -1.641 | 0 | 0.263 |
| 75 | AC003991.3 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-142-3p;hsa-miR-17-5p;hsa-miR-193a-3p;hsa-miR-193b-3p;hsa-miR-19b-1-5p;hsa-miR-20a-5p;hsa-miR-20b-5p;hsa-miR-365a-3p;hsa-miR-425-5p;hsa-miR-769-3p;hsa-miR-92a-3p;hsa-miR-93-5p | 14 | CCND1 | Sponge network | -0.787 | 0.08132 | -0.296 | 0.2554 | 0.255 |
| 76 | RP11-53M11.3 | hsa-miR-130b-5p;hsa-miR-141-3p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-191-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-301a-3p;hsa-miR-7-1-3p | 10 | CCND2 | Sponge network | -2.058 | 0.01204 | -1.641 | 0 | 0.253 |