This regulatory network was inferred from the input dataset. The miRNAs and mRNAs are
presented as round and rectangle nodes respectively. The numerical value popped up upon mouse over the gene node is the log2 transformed fold-change of the gene expression between the two groups. All of the nodes are clickable, and the detailed information of the miRNAs/mRNAs and related cancer pathway will be displayed in another window. The edges between nodes are supported by both interactions (predicted or experimentally verified) and correlations learnt from cancer dataset. The numerical value popped up upon mouse over the edge is the correlation beat value (effect size) between the two nodes. The experimental evidences of the edges reported in previous cancer studies are highlighted by red/orange color. All of these information can be accessed by the "mouse-over" action. This network shows a full map of the miRNA-mRNA regulation of the input gene list(s), and the hub miRNAs (with the high network degree/betweenness centrality) would be the potential cancer drivers or tumor suppressors. The full result table can be accessed in the "Regulations" tab.
"miRNACancerMAP" is also a network visualization tool for users to draw their regulatory network by personal customization. Users can set the complexity of the network by limiting the number of nodes or edges. And the color of the nodes can be defined by different categories of the mRNAs and miRNAs, such as Gene-Ontology, pathway, and expression status. Users can also select to use network degree or network betweenness centrality to define the node size. And edges can be black or colored by the correlation. Purple edge means negative correlation (mostly found between miRNA and mRNA), and blue edge means positive correlation (found in PPI or miRNA-miRNA sponge effect). We can also add the protein-protein interactions (PPI) into the network. This result will show the cluster of genes regulated by some specific miRNAs. Additionally, miRNA-miRNA edges can be added by the "miRNA sponge" button, presenting some clusters of miRNAs that have the interactions via sponge effect.
miRNA-gene regulations
| Num | microRNA | Gene | miRNA log2FC | miRNA pvalue | Gene log2FC | Gene pvalue | Interaction | Correlation beta | Correlation P-value | PMID | Reported in cancer studies |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | hsa-miR-106a-5p | BMP2 | 1.39 | 6.0E-5 | -2.2 | 0 | miRNATAP | -0.25 | 0 | NA | |
| 2 | hsa-miR-106b-5p | BMP2 | 1.47 | 0 | -2.2 | 0 | miRNATAP | -0.32 | 7.0E-5 | NA | |
| 3 | hsa-miR-142-5p | BMP2 | 1.3 | 0 | -2.2 | 0 | PITA; miRNATAP | -0.19 | 0.00333 | NA | |
| 4 | hsa-miR-17-5p | BMP2 | 2.07 | 0 | -2.2 | 0 | TargetScan | -0.28 | 5.0E-5 | NA | |
| 5 | hsa-miR-20a-5p | BMP2 | 2.65 | 0 | -2.2 | 0 | miRNATAP | -0.24 | 0.00012 | NA | |
| 6 | hsa-miR-335-3p | BMP2 | 1.51 | 0 | -2.2 | 0 | mirMAP | -0.26 | 0.00065 | NA | |
| 7 | hsa-miR-362-3p | BMP2 | 0.19 | 0.52808 | -2.2 | 0 | miRanda | -0.14 | 0.04028 | NA | |
| 8 | hsa-miR-421 | BMP2 | 0.17 | 0.53528 | -2.2 | 0 | PITA; miRanda | -0.26 | 9.0E-5 | NA | |
| 9 | hsa-miR-590-3p | BMP2 | 0.84 | 0.00129 | -2.2 | 0 | PITA; miRanda; mirMAP | -0.31 | 5.0E-5 | NA | |
| 10 | hsa-miR-106b-5p | BMP7 | 1.47 | 0 | -1 | 0.05546 | mirMAP | -0.45 | 0.00015 | NA | |
| 11 | hsa-miR-130b-5p | BMP7 | 1.54 | 0 | -1 | 0.05546 | mirMAP | -0.43 | 0 | NA | |
| 12 | hsa-miR-17-5p | BMP7 | 2.07 | 0 | -1 | 0.05546 | mirMAP | -0.32 | 0.0013 | NA | |
| 13 | hsa-miR-20a-5p | BMP7 | 2.65 | 0 | -1 | 0.05546 | mirMAP | -0.2 | 0.02795 | NA | |
| 14 | hsa-miR-93-5p | BMP7 | 1.51 | 0 | -1 | 0.05546 | mirMAP | -0.27 | 0.01283 | NA | |
| 15 | hsa-miR-532-3p | CTNNB1 | -0.68 | 0.01476 | -0.01 | 0.94149 | miRNAWalker2 validate | -0.11 | 1.0E-5 | NA | |
| 16 | hsa-miR-134-5p | EPB41L5 | 0.17 | 0.62286 | -1.18 | 0 | MirTarget | -0.13 | 0 | NA | |
| 17 | hsa-miR-148a-3p | EPB41L5 | 2.31 | 0 | -1.18 | 0 | MirTarget | -0.18 | 0 | NA | |
| 18 | hsa-miR-148a-5p | EPB41L5 | 1.46 | 0 | -1.18 | 0 | mirMAP | -0.14 | 4.0E-5 | NA | |
| 19 | hsa-miR-152-3p | EPB41L5 | 0.66 | 0.0013 | -1.18 | 0 | MirTarget | -0.26 | 0 | NA | |
| 20 | hsa-miR-424-5p | EPB41L5 | 1.26 | 1.0E-5 | -1.18 | 0 | MirTarget | -0.12 | 7.0E-5 | NA | |
| 21 | hsa-miR-590-5p | EPB41L5 | 2.07 | 0 | -1.18 | 0 | miRanda | -0.15 | 9.0E-5 | NA | |
| 22 | hsa-miR-139-5p | FAM83D | -2.27 | 0 | 2.05 | 0 | miRanda | -0.4 | 0 | NA | |
| 23 | hsa-miR-3065-5p | FAM83D | 0.65 | 0.09995 | 2.05 | 0 | mirMAP | -0.29 | 0 | NA | |
| 24 | hsa-miR-486-5p | FAM83D | -4.39 | 0 | 2.05 | 0 | miRanda | -0.21 | 0 | NA | |
| 25 | hsa-miR-146b-5p | FGFR2 | 1.09 | 1.0E-5 | -2.39 | 0 | miRanda | -0.18 | 0.00873 | NA | |
| 26 | hsa-miR-186-5p | FGFR2 | 0.85 | 0 | -2.39 | 0 | miRNAWalker2 validate | -0.24 | 0.04721 | NA | |
| 27 | hsa-miR-19b-1-5p | FGFR2 | 1.71 | 0 | -2.39 | 0 | miRNAWalker2 validate; miRTarBase | -0.27 | 0.00061 | NA | |
| 28 | hsa-miR-324-5p | FGFR2 | 1.07 | 5.0E-5 | -2.39 | 0 | miRanda | -0.15 | 0.02348 | NA | |
| 29 | hsa-miR-542-3p | FGFR2 | 1.62 | 0 | -2.39 | 0 | miRanda | -0.19 | 0.00349 | NA | |
| 30 | hsa-miR-590-3p | FGFR2 | 0.84 | 0.00129 | -2.39 | 0 | miRanda | -0.38 | 0 | NA | |
| 31 | hsa-miR-130b-3p | FOXF2 | 1.83 | 0 | -1.7 | 0 | miRNATAP | -0.54 | 0 | NA | |
| 32 | hsa-miR-142-3p | FOXF2 | 3.98 | 0 | -1.7 | 0 | miRanda | -0.14 | 0.00021 | NA | |
| 33 | hsa-miR-182-5p | FOXF2 | 3.22 | 0 | -1.7 | 0 | miRNAWalker2 validate; miRNATAP | -0.37 | 0 | 25738520 | miR 182 promotes cell growth and invasion by targeting forkhead box F2 transcription factor in colorectal cancer; In the present study we aimed to investigate the role of miR-182 in colorectal cancer CRC and identify the regulation of FoxF2 by miR-182; The direct binding of miR-182 to the 3' untranslated region 3'UTR of FoxF2 mRNA was confirmed using a luciferase reporter gene assay |
| 34 | hsa-miR-19a-3p | FOXF2 | 2.12 | 0 | -1.7 | 0 | miRNATAP | -0.35 | 0 | NA | |
| 35 | hsa-miR-19b-3p | FOXF2 | 2.11 | 0 | -1.7 | 0 | miRNATAP | -0.35 | 0 | NA | |
| 36 | hsa-miR-200c-3p | FOXF2 | 0.38 | 0.08422 | -1.7 | 0 | miRNATAP | -0.27 | 1.0E-5 | NA | |
| 37 | hsa-miR-301a-3p | FOXF2 | 2.7 | 0 | -1.7 | 0 | miRNATAP | -0.32 | 0 | NA | |
| 38 | hsa-miR-429 | FOXF2 | 2.38 | 0 | -1.7 | 0 | PITA; miRanda; miRNATAP | -0.13 | 0.00296 | NA | |
| 39 | hsa-miR-454-3p | FOXF2 | 1.49 | 0 | -1.7 | 0 | miRNATAP | -0.42 | 0 | NA | |
| 40 | hsa-miR-590-3p | FOXF2 | 0.84 | 0.00129 | -1.7 | 0 | PITA; miRanda | -0.33 | 0 | NA | |
| 41 | hsa-miR-9-5p | FOXF2 | 4.99 | 0 | -1.7 | 0 | miRNATAP | -0.17 | 0 | NA | |
| 42 | hsa-miR-96-5p | FOXF2 | 3.04 | 0 | -1.7 | 0 | TargetScan; miRNATAP | -0.42 | 0 | NA | |
| 43 | hsa-miR-146b-5p | GSK3B | 1.09 | 1.0E-5 | -0.32 | 0.0051 | miRanda | -0.12 | 0 | NA | |
| 44 | hsa-miR-181a-5p | GSK3B | -0.38 | 0.05621 | -0.32 | 0.0051 | mirMAP | -0.13 | 0 | NA | |
| 45 | hsa-miR-181b-5p | GSK3B | 0.67 | 0.00024 | -0.32 | 0.0051 | mirMAP | -0.14 | 0 | NA | |
| 46 | hsa-miR-199a-5p | GSK3B | 1.31 | 0 | -0.32 | 0.0051 | PITA; miRanda; miRNATAP | -0.12 | 0 | NA | |
| 47 | hsa-miR-199b-5p | GSK3B | 2.14 | 0 | -0.32 | 0.0051 | PITA; miRanda; miRNATAP | -0.1 | 0 | NA | |
| 48 | hsa-miR-26b-5p | GSK3B | 0.72 | 5.0E-5 | -0.32 | 0.0051 | miRNATAP | -0.16 | 0 | NA | |
| 49 | hsa-miR-29a-3p | GSK3B | 0.1 | 0.5732 | -0.32 | 0.0051 | miRNATAP | -0.12 | 7.0E-5 | NA | |
| 50 | hsa-miR-29b-3p | GSK3B | 3.11 | 0 | -0.32 | 0.0051 | miRTarBase; miRNATAP | -0.12 | 0 | NA | |
| 51 | hsa-miR-342-3p | GSK3B | -0.13 | 0.56103 | -0.32 | 0.0051 | miRanda | -0.12 | 0 | NA | |
| 52 | hsa-miR-664a-3p | GSK3B | 0.44 | 0.02142 | -0.32 | 0.0051 | mirMAP | -0.14 | 0 | NA | |
| 53 | hsa-let-7a-3p | HGF | 0.17 | 0.43183 | -1.47 | 2.0E-5 | mirMAP | -0.22 | 0.00866 | NA | |
| 54 | hsa-let-7f-1-3p | HGF | -0.09 | 0.69397 | -1.47 | 2.0E-5 | mirMAP | -0.21 | 0.01316 | NA | |
| 55 | hsa-miR-141-3p | HGF | 3.37 | 0 | -1.47 | 2.0E-5 | MirTarget; TargetScan | -0.4 | 0 | NA | |
| 56 | hsa-miR-200a-3p | HGF | 3.15 | 0 | -1.47 | 2.0E-5 | MirTarget | -0.25 | 0 | NA | |
| 57 | hsa-miR-203a-3p | HGF | 1.07 | 0.0039 | -1.47 | 2.0E-5 | MirTarget | -0.15 | 0.0003 | NA | |
| 58 | hsa-miR-26b-5p | HGF | 0.72 | 5.0E-5 | -1.47 | 2.0E-5 | MirTarget; miRNATAP | -0.31 | 0.00059 | NA | |
| 59 | hsa-miR-29a-5p | HGF | 1.9 | 0 | -1.47 | 2.0E-5 | mirMAP | -0.29 | 1.0E-5 | NA | |
| 60 | hsa-miR-30b-5p | HGF | 0.36 | 0.13803 | -1.47 | 2.0E-5 | mirMAP | -0.23 | 0.00049 | NA | |
| 61 | hsa-miR-335-3p | HGF | 1.51 | 0 | -1.47 | 2.0E-5 | mirMAP | -0.18 | 0.01397 | NA | |
| 62 | hsa-miR-421 | HGF | 0.17 | 0.53528 | -1.47 | 2.0E-5 | miRanda | -0.19 | 0.00222 | NA | |
| 63 | hsa-miR-429 | HGF | 2.38 | 0 | -1.47 | 2.0E-5 | miRanda | -0.33 | 0 | NA | |
| 64 | hsa-miR-542-3p | HGF | 1.62 | 0 | -1.47 | 2.0E-5 | miRanda | -0.19 | 0.00164 | NA | |
| 65 | hsa-miR-590-3p | HGF | 0.84 | 0.00129 | -1.47 | 2.0E-5 | miRanda; mirMAP | -0.35 | 0 | NA | |
| 66 | hsa-miR-590-5p | HGF | 2.07 | 0 | -1.47 | 2.0E-5 | miRanda | -0.41 | 0 | NA | |
| 67 | hsa-miR-651-5p | HGF | 1.67 | 0 | -1.47 | 2.0E-5 | MirTarget | -0.28 | 0 | NA | |
| 68 | hsa-miR-7-1-3p | HGF | 2.61 | 0 | -1.47 | 2.0E-5 | mirMAP | -0.29 | 5.0E-5 | NA | |
| 69 | hsa-miR-139-5p | HIF1A | -2.27 | 0 | 0.42 | 0.01526 | miRanda | -0.12 | 1.0E-5 | NA | |
| 70 | hsa-miR-195-3p | HIF1A | -1.33 | 0 | 0.42 | 0.01526 | MirTarget | -0.14 | 6.0E-5 | NA | |
| 71 | hsa-miR-320a | HIF1A | -0.96 | 0 | 0.42 | 0.01526 | miRanda | -0.13 | 0.00665 | NA | |
| 72 | hsa-miR-660-5p | HIF1A | 2.05 | 0 | 0.42 | 0.01526 | MirTarget | -0.16 | 0 | NA | |
| 73 | hsa-let-7a-5p | HMGA2 | -1.37 | 0 | 3.12 | 0.00011 | miRNAWalker2 validate; miRTarBase; MirTarget; TargetScan | -0.44 | 0.03109 | 23700794; 20949044; 18413822; 23073586; 21598109; 24612219; 23134218; 21412931 | MiR-98 a member in the let-7 family acts as a negative regulator in the expression of HMGA2 high mobility group A2 oncogene and it has been shown to have a nearly 3-fold decrease in A549/DDP cells;Since let-7 miRNAs generally play a tumor-suppressive role as targeting oncogenes such as RAS and HMGA2 our results suggest that AZ-P7a cells release let-7 miRNAs via exosomes into the extracellular environment to maintain their oncogenesis;Recent studies report that HMGA2 is negatively regulated by the let-7 microRNA miRNA family; However no studies have examined the clinical significance of HMGA2 and its relationship to the let-7 miRNA family in gastric cancer; We also did an association study comparing HMGA2 expression and let-7 miRNA family expression in gastric cancer; An inverse correlation between HMGA2 and let-7a was found in gastric cancer cell lines P = 0.08; Furthermore our findings suggest that HMGA2 is negatively regulated by the let-7 miRNA family in human gastric cancer;HMGA2 mRNA and let-7 family microRNA were detected by real time fluorescent quantitative reverse transcription polymerase chain reaction in the corresponding frozen tissues; All let-7 family members were detectable in all ovarian cancer samples and their expression were inversely correlated with HMGA2 mRNA expression r=-0.305P<0.05; The downregulation of let-7 is but not the only mechanism of HMGA2 overexpression in serous ovarian cancer;Role of microRNA let 7 and effect to HMGA2 in esophageal squamous cell carcinoma; To investigated the role of microRNA miRNA let-7 and its regulation on high mobility group A2 HMGA2 protein expression in esophageal squamous cell carcinoma ESCC; To evaluate the role of let-7 and HMGA2 cell proliferations were analyzed with synthetic let-7 mimics- or its inhibitor-transfected cells; The transcription of let-7 inversely correlated with HMGA2 protein; The present results demonstrated that let-7 and HMGA2 involved in ESCC carcinogenesis;HMGA2 is down regulated by microRNA let 7 and associated with epithelial mesenchymal transition in oesophageal squamous cell carcinomas of Kazakhs; To investigate the expression of let-7 and its regulation of high-mobility group A2 protein HMGA2 and to verify the relationship between let-7 HMGA2 and the process of epithelial-mesenchymal transition EMT in oesophageal squamous cell carcinomas OSCC of Kazakh patients; Let-7 could repress expression of HMGA2 after co-transfection with let-7 and HMGA2 P = 0.002; Moreover let-7 expression was observed less frequently P = 2.0 × 10-8 and HMGA2 expression more frequently P = 1.0 × 10-10 in OSCC than in normal adjacent tissues; and let-7 expression was observed less frequently in OSCC from Kazakh patients than in those from Han and Uygur patients P = 0.041; There was a reverse correlation between expression of let-7 and HMGA2 P = 0.018; Expression of let-7 can suppress cell proliferation by acting directly on regulation of HMGA2 in OSCC;MicroRNA let 7a inhibits the proliferation and invasion of nonsmall cell lung cancer cell line 95D by regulating K Ras and HMGA2 gene expression; K-RAS and HMGA2 mRNA levels were significantly higher in the let-7a overexpressed group than those in the let-7a inhibited group p < 0.05; However the protein levels of K-RAS and HMGA2 were significantly lower in the let-7a overexpressed group than those in the let-7a inhibited group p < 0.05; We suppose that let-7a inhibits the proliferation and invasion of the cell line 95D by regulating the translation of K-RAS and HMGA2 mRNA not the transcription of the mRNA itself;Let 7 microRNA and HMGA2 levels of expression are not inversely linked in adipocytic tumors: analysis of 56 lipomas and liposarcomas with molecular cytogenetic data; The aim of our study was first to assess the role of HMGA2 expression in the pathogenesis of adipocytic tumors AT and second to seek a potential correlation between overexpression of HMGA2 and let-7 expression inhibition by analyzing a series of 56 benign and malignant AT with molecular cytogenetic data; We measured the levels of expression of HMGA2 mRNA and of eight members of the let-7 microRNA family using quantitative RT-PCR and expression of HMGA2 protein using immunohistochemistry; Although overexpression of both HMGA2 mRNA and protein in a majority of ordinary lipomas without HMGA2 structural rearrangement may have suggested a potential role for let-7 microRNAs we did not observe a significant link with let-7 inhibition in such cases |
| 74 | hsa-let-7b-5p | HMGA2 | -1.62 | 0 | 3.12 | 0.00011 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.36 | 0.04256 | 23700794; 20949044; 18413822; 23073586; 21598109; 24612219; 21412931 | MiR-98 a member in the let-7 family acts as a negative regulator in the expression of HMGA2 high mobility group A2 oncogene and it has been shown to have a nearly 3-fold decrease in A549/DDP cells;Since let-7 miRNAs generally play a tumor-suppressive role as targeting oncogenes such as RAS and HMGA2 our results suggest that AZ-P7a cells release let-7 miRNAs via exosomes into the extracellular environment to maintain their oncogenesis;Recent studies report that HMGA2 is negatively regulated by the let-7 microRNA miRNA family; However no studies have examined the clinical significance of HMGA2 and its relationship to the let-7 miRNA family in gastric cancer; We also did an association study comparing HMGA2 expression and let-7 miRNA family expression in gastric cancer; Furthermore our findings suggest that HMGA2 is negatively regulated by the let-7 miRNA family in human gastric cancer;HMGA2 mRNA and let-7 family microRNA were detected by real time fluorescent quantitative reverse transcription polymerase chain reaction in the corresponding frozen tissues; All let-7 family members were detectable in all ovarian cancer samples and their expression were inversely correlated with HMGA2 mRNA expression r=-0.305P<0.05; The downregulation of let-7 is but not the only mechanism of HMGA2 overexpression in serous ovarian cancer;Role of microRNA let 7 and effect to HMGA2 in esophageal squamous cell carcinoma; To investigated the role of microRNA miRNA let-7 and its regulation on high mobility group A2 HMGA2 protein expression in esophageal squamous cell carcinoma ESCC; To evaluate the role of let-7 and HMGA2 cell proliferations were analyzed with synthetic let-7 mimics- or its inhibitor-transfected cells; The transcription of let-7 inversely correlated with HMGA2 protein; The present results demonstrated that let-7 and HMGA2 involved in ESCC carcinogenesis;HMGA2 is down regulated by microRNA let 7 and associated with epithelial mesenchymal transition in oesophageal squamous cell carcinomas of Kazakhs; To investigate the expression of let-7 and its regulation of high-mobility group A2 protein HMGA2 and to verify the relationship between let-7 HMGA2 and the process of epithelial-mesenchymal transition EMT in oesophageal squamous cell carcinomas OSCC of Kazakh patients; Let-7 could repress expression of HMGA2 after co-transfection with let-7 and HMGA2 P = 0.002; Moreover let-7 expression was observed less frequently P = 2.0 × 10-8 and HMGA2 expression more frequently P = 1.0 × 10-10 in OSCC than in normal adjacent tissues; and let-7 expression was observed less frequently in OSCC from Kazakh patients than in those from Han and Uygur patients P = 0.041; There was a reverse correlation between expression of let-7 and HMGA2 P = 0.018; Expression of let-7 can suppress cell proliferation by acting directly on regulation of HMGA2 in OSCC;Let 7 microRNA and HMGA2 levels of expression are not inversely linked in adipocytic tumors: analysis of 56 lipomas and liposarcomas with molecular cytogenetic data; The aim of our study was first to assess the role of HMGA2 expression in the pathogenesis of adipocytic tumors AT and second to seek a potential correlation between overexpression of HMGA2 and let-7 expression inhibition by analyzing a series of 56 benign and malignant AT with molecular cytogenetic data; We measured the levels of expression of HMGA2 mRNA and of eight members of the let-7 microRNA family using quantitative RT-PCR and expression of HMGA2 protein using immunohistochemistry; Although overexpression of both HMGA2 mRNA and protein in a majority of ordinary lipomas without HMGA2 structural rearrangement may have suggested a potential role for let-7 microRNAs we did not observe a significant link with let-7 inhibition in such cases |
| 75 | hsa-let-7c-5p | HMGA2 | -2.14 | 0 | 3.12 | 0.00011 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.32 | 0.03136 | NA | |
| 76 | hsa-let-7d-5p | HMGA2 | -0.62 | 6.0E-5 | 3.12 | 0.00011 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.65 | 0.00736 | NA | |
| 77 | hsa-let-7g-5p | HMGA2 | 0.08 | 0.58308 | 3.12 | 0.00011 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.58 | 0.01931 | 21472347; 18308936 | Furthermore K-Ras and HMGA2 are well known as targets of let-7g; In this study we evaluated the potential role of precursor pre-let-7g in lung cancer cell metastasis focusing on the two targets of let-7g HMGA2 and K-Ras;In let-7g expressing tumors reductions in Ras family and HMGA2 protein levels were detected; Ectopic expression of K-RasG12D largely rescued let-7g mediated tumor suppression whereas ectopic expression of HMGA2 was less effective |
| 78 | hsa-miR-129-5p | HMGA2 | -0.41 | 0.34149 | 3.12 | 0.00011 | miRanda; mirMAP | -0.2 | 0.02051 | NA | |
| 79 | hsa-miR-195-3p | HMGA2 | -1.33 | 0 | 3.12 | 0.00011 | mirMAP | -0.43 | 0.0073 | NA | |
| 80 | hsa-miR-195-5p | HMGA2 | -1.02 | 5.0E-5 | 3.12 | 0.00011 | MirTarget | -0.45 | 0.00378 | NA | |
| 81 | hsa-miR-204-5p | HMGA2 | -1.02 | 0.00767 | 3.12 | 0.00011 | miRNATAP | -0.28 | 0.005 | 27095441; 26406941 | We identified high mobility group protein A2 HMGA2 as a novel direct target of miR-204 and showed that miR-204 expression was decreased while HMGA2 expression was increased in CRC cell lines; Additionally both MiR-204 overexpression and HMGA2 inhibition attenuated cell proliferation whereas forced expression of HMGA2 partly restored the inhibitory effect of miR-204 on HCT116 and SW480 cells; Taken together our present study elucidated that miR-204 upregulated 5-Fu chemosensitivity via the downregulation of HMGA2 in colorectal cancer and provided significant insight into the mechanism of 5-Fu resistance in colorectal cancer patients;MiR 204 regulates HMGA2 expression and inhibits cell proliferation in human thyroid cancer; Analysis using publicly available algorithms has found that high mobility group AT-hook 2 HMGA2 a key transcriptional regulatory factor is a potential target of microRNA-204 miR-204; Some studies have shown that both miR-204 and HMGA2 are associated with cancer development; We examined the possible relationship between miR-204 and HMGA2 in the development of thyroid cancer; MiR-204 was downregulated in the thyroid cancer specimens and cell lines and targeted the 3^\prime untranslated region of HMGA2 directly |
| 82 | hsa-miR-26b-5p | HMGA2 | 0.72 | 5.0E-5 | 3.12 | 0.00011 | miRNATAP | -0.46 | 0.02837 | NA | |
| 83 | hsa-miR-29b-2-5p | HMGA2 | 0.35 | 0.19484 | 3.12 | 0.00011 | mirMAP | -0.69 | 0 | NA | |
| 84 | hsa-miR-3065-5p | HMGA2 | 0.65 | 0.09995 | 3.12 | 0.00011 | mirMAP; miRNATAP | -0.25 | 0.01661 | NA | |
| 85 | hsa-miR-30b-5p | HMGA2 | 0.36 | 0.13803 | 3.12 | 0.00011 | mirMAP | -0.66 | 2.0E-5 | NA | |
| 86 | hsa-miR-30c-5p | HMGA2 | -0.33 | 0.1236 | 3.12 | 0.00011 | mirMAP | -0.44 | 0.01213 | NA | |
| 87 | hsa-miR-30d-3p | HMGA2 | 0 | 0.98646 | 3.12 | 0.00011 | MirTarget | -0.83 | 0 | NA | |
| 88 | hsa-miR-30d-5p | HMGA2 | -0.92 | 4.0E-5 | 3.12 | 0.00011 | mirMAP | -1.03 | 0 | NA | |
| 89 | hsa-miR-32-5p | HMGA2 | 0.88 | 6.0E-5 | 3.12 | 0.00011 | miRNATAP | -0.51 | 0.00603 | NA | |
| 90 | hsa-miR-326 | HMGA2 | -0.99 | 0.00335 | 3.12 | 0.00011 | miRNATAP | -0.42 | 0.00013 | NA | |
| 91 | hsa-miR-362-3p | HMGA2 | 0.19 | 0.52808 | 3.12 | 0.00011 | miRanda | -0.32 | 0.03491 | NA | |
| 92 | hsa-miR-486-5p | HMGA2 | -4.39 | 0 | 3.12 | 0.00011 | miRanda | -0.24 | 0.00642 | NA | |
| 93 | hsa-miR-491-5p | HMGA2 | -0.47 | 0.06833 | 3.12 | 0.00011 | PITA; miRanda | -0.35 | 0.01695 | NA | |
| 94 | hsa-miR-664a-3p | HMGA2 | 0.44 | 0.02142 | 3.12 | 0.00011 | mirMAP | -0.93 | 0 | NA | |
| 95 | hsa-miR-664a-5p | HMGA2 | -0.09 | 0.66227 | 3.12 | 0.00011 | MirTarget | -0.73 | 5.0E-5 | NA | |
| 96 | hsa-miR-139-5p | HNRNPAB | -2.27 | 0 | 0.65 | 0 | miRanda | -0.12 | 0 | NA | |
| 97 | hsa-miR-107 | LEF1 | 0.66 | 0 | 0.59 | 0.00774 | miRanda | -0.26 | 0.00074 | NA | |
| 98 | hsa-miR-26b-5p | LEF1 | 0.72 | 5.0E-5 | 0.59 | 0.00774 | miRNATAP | -0.14 | 0.015 | 24785257 | miR26b is downregulated in several cancers and tumors and miR26b directly targets the lymphoid enhancer factor 1 Lef13'UTR and inhibits endogenous Lef1 expression; We report that miR26b expression is associated with human colon cancer through the regulation of LEF1 expression in colon cancer cells; Analyses of multiple colon cancer cell lines revealed an inverse correlation between miR26b and LEF1 expression; Normal human colon cells express low levels of LEF1 and high levels of miR26b; however human colon cancer cells have decreased miR26b expression and increased LEF1 expression; We demonstrate that miR26b expression is a potent inhibitor of colon cancer cell proliferation and significantly decreases LEF1 expression; Analyses of human colon cancer databases also demonstrated a link between miR26b and LEF1 expression |
| 99 | hsa-miR-3065-3p | LEF1 | 0.38 | 0.30796 | 0.59 | 0.00774 | miRNATAP | -0.12 | 1.0E-5 | NA | |
| 100 | hsa-miR-3065-5p | LEF1 | 0.65 | 0.09995 | 0.59 | 0.00774 | miRNATAP | -0.13 | 0 | NA | |
| 101 | hsa-miR-335-3p | LEF1 | 1.51 | 0 | 0.59 | 0.00774 | mirMAP | -0.11 | 0.02649 | NA | |
| 102 | hsa-let-7a-3p | LIMS1 | 0.17 | 0.43183 | -0.7 | 0.00015 | mirMAP | -0.15 | 0.00066 | NA | |
| 103 | hsa-miR-148b-5p | LIMS1 | 1.39 | 0 | -0.7 | 0.00015 | MirTarget | -0.21 | 0 | NA | |
| 104 | hsa-miR-182-5p | LIMS1 | 3.22 | 0 | -0.7 | 0.00015 | MirTarget; miRNATAP | -0.12 | 0.00027 | NA | |
| 105 | hsa-miR-186-5p | LIMS1 | 0.85 | 0 | -0.7 | 0.00015 | mirMAP | -0.23 | 0.00018 | NA | |
| 106 | hsa-miR-29a-3p | LIMS1 | 0.1 | 0.5732 | -0.7 | 0.00015 | MirTarget; miRNATAP | -0.18 | 0.00022 | NA | |
| 107 | hsa-miR-29b-3p | LIMS1 | 3.11 | 0 | -0.7 | 0.00015 | MirTarget; miRNATAP | -0.25 | 0 | NA | |
| 108 | hsa-miR-30b-5p | LIMS1 | 0.36 | 0.13803 | -0.7 | 0.00015 | MirTarget | -0.32 | 0 | NA | |
| 109 | hsa-miR-30c-5p | LIMS1 | -0.33 | 0.1236 | -0.7 | 0.00015 | MirTarget | -0.28 | 0 | NA | |
| 110 | hsa-miR-30d-3p | LIMS1 | 0 | 0.98646 | -0.7 | 0.00015 | mirMAP | -0.16 | 8.0E-5 | NA | |
| 111 | hsa-miR-30d-5p | LIMS1 | -0.92 | 4.0E-5 | -0.7 | 0.00015 | MirTarget | -0.14 | 0.00037 | NA | |
| 112 | hsa-miR-30e-5p | LIMS1 | 1.6 | 0 | -0.7 | 0.00015 | MirTarget | -0.17 | 0.00019 | NA | |
| 113 | hsa-miR-429 | LIMS1 | 2.38 | 0 | -0.7 | 0.00015 | miRanda; miRNATAP | -0.22 | 0 | NA | |
| 114 | hsa-miR-7-1-3p | LIMS1 | 2.61 | 0 | -0.7 | 0.00015 | mirMAP | -0.11 | 0.00311 | NA | |
| 115 | hsa-miR-96-5p | LIMS1 | 3.04 | 0 | -0.7 | 0.00015 | TargetScan; miRNATAP | -0.23 | 0 | NA | |
| 116 | hsa-miR-338-3p | LOXL2 | 0.73 | 0.05063 | 1.02 | 0.00103 | miRanda | -0.18 | 0 | NA | |
| 117 | hsa-let-7a-5p | LOXL3 | -1.37 | 0 | -0.11 | 0.58907 | miRNATAP | -0.19 | 0.00024 | NA | |
| 118 | hsa-let-7d-5p | LOXL3 | -0.62 | 6.0E-5 | -0.11 | 0.58907 | miRNATAP | -0.19 | 0.00168 | NA | |
| 119 | hsa-let-7e-5p | LOXL3 | -0.75 | 7.0E-5 | -0.11 | 0.58907 | miRNATAP | -0.11 | 0.02879 | NA | |
| 120 | hsa-let-7f-5p | LOXL3 | -0.05 | 0.83408 | -0.11 | 0.58907 | miRNATAP | -0.17 | 1.0E-5 | NA | |
| 121 | hsa-let-7g-5p | LOXL3 | 0.08 | 0.58308 | -0.11 | 0.58907 | miRNATAP | -0.19 | 0.00289 | NA | |
| 122 | hsa-miR-2110 | LOXL3 | -1.92 | 0 | -0.11 | 0.58907 | miRNATAP | -0.14 | 4.0E-5 | NA | |
| 123 | hsa-miR-142-5p | NOG | 1.3 | 0 | -0.7 | 0.1118 | miRNATAP | -0.18 | 0.01861 | NA | |
| 124 | hsa-miR-186-5p | NOG | 0.85 | 0 | -0.7 | 0.1118 | MirTarget; miRNATAP | -0.36 | 0.01474 | NA | |
| 125 | hsa-miR-200c-3p | NOG | 0.38 | 0.08422 | -0.7 | 0.1118 | MirTarget; miRNATAP | -0.24 | 0.00944 | NA | |
| 126 | hsa-miR-28-5p | NOTCH1 | 1.2 | 0 | -1.08 | 0 | miRanda | -0.24 | 7.0E-5 | NA | |
| 127 | hsa-miR-664a-3p | RBPJ | 0.44 | 0.02142 | 0.07 | 0.55019 | mirMAP | -0.13 | 2.0E-5 | NA | |
| 128 | hsa-let-7a-3p | SNAI1 | 0.17 | 0.43183 | -0.76 | 0.00279 | miRNATAP | -0.15 | 0.01905 | NA | |
| 129 | hsa-miR-26b-5p | SNAI1 | 0.72 | 5.0E-5 | -0.76 | 0.00279 | miRNAWalker2 validate | -0.16 | 0.01278 | NA | |
| 130 | hsa-miR-30a-5p | SNAI1 | -0.92 | 0.00076 | -0.76 | 0.00279 | miRTarBase; miRNATAP | -0.14 | 0.00089 | 24954667; 27212164; 21633953 | We identified SNAI1 as a direct target of miR-30a and demonstrated miR-30a as a novel regulator of EMT by targeting SNAI1 indicating its potential therapeutic value for reducing invasion and metastasis of HCC;Knockdown of endogenous miR-30a promoted the elongated fibroblast-like morphologic alteration of SGC-7901 cells and also enhanced Snail and Vimentin expression; MiR-30a overexpression induced morphological changes from an extended fibroblast-like morphology to more epithelial-like morphology in SGC-7901/DDP cells and decreased Snail and Vimentin level;MicroRNA 30a inhibits epithelial to mesenchymal transition by targeting Snai1 and is downregulated in non small cell lung cancer; Forced re-introduction of miR-30a significantly altered cell morphology in vitro invasion and migration of invasive cell lines this being paralleled by a downregulation of Snai1 and upregulation of E-cadherin expression; These results suggest that miR-30a targets Snai1 inhibits invasion and metastasis and is downregulated in NSCLC |
| 131 | hsa-miR-30b-5p | SNAI1 | 0.36 | 0.13803 | -0.76 | 0.00279 | miRTarBase; miRNATAP | -0.29 | 0 | NA | |
| 132 | hsa-miR-30c-5p | SNAI1 | -0.33 | 0.1236 | -0.76 | 0.00279 | miRTarBase; miRNATAP | -0.24 | 1.0E-5 | NA | |
| 133 | hsa-miR-30d-5p | SNAI1 | -0.92 | 4.0E-5 | -0.76 | 0.00279 | miRTarBase; miRNATAP | -0.4 | 0 | 26501435 | miR 30d Blocked Transforming Growth Factor β1 Induced Epithelial Mesenchymal Transition by Targeting Snail in Ovarian Cancer Cells; Luciferase activity assay was performed to verify the direct inhibition of Snail by miR-30d; Furthermore Snail was identified as the direct target of miR-30d; Our results revealed that miR-30d functioned as a suppressor of ovarian cancer progression by decreasing Snail expression and thus blocking TGF-β1-induced EMT process suggesting the potentiality of miR-30d analogs to be used as therapeutics for ovarian cancer |
| 134 | hsa-miR-30e-5p | SNAI1 | 1.6 | 0 | -0.76 | 0.00279 | miRTarBase | -0.3 | 0 | NA | |
| 135 | hsa-miR-335-5p | SNAI1 | -0.47 | 0.0677 | -0.76 | 0.00279 | miRNAWalker2 validate | -0.16 | 0.0019 | NA | |
| 136 | hsa-miR-107 | SNAI2 | 0.66 | 0 | -0.55 | 0.05725 | miRanda | -0.45 | 1.0E-5 | NA | |
| 137 | hsa-miR-148b-3p | SNAI2 | 0.48 | 0.00265 | -0.55 | 0.05725 | miRNAWalker2 validate | -0.56 | 0 | NA | |
| 138 | hsa-miR-181a-5p | SNAI2 | -0.38 | 0.05621 | -0.55 | 0.05725 | miRNAWalker2 validate | -0.14 | 0.04285 | NA | |
| 139 | hsa-miR-182-5p | SNAI2 | 3.22 | 0 | -0.55 | 0.05725 | miRNAWalker2 validate; miRNATAP | -0.28 | 0 | NA | |
| 140 | hsa-miR-200b-3p | SNAI2 | 1.55 | 0 | -0.55 | 0.05725 | TargetScan | -0.44 | 0 | NA | |
| 141 | hsa-miR-200c-3p | SNAI2 | 0.38 | 0.08422 | -0.55 | 0.05725 | miRNATAP | -0.45 | 0 | NA | |
| 142 | hsa-miR-335-3p | SNAI2 | 1.51 | 0 | -0.55 | 0.05725 | mirMAP | -0.28 | 1.0E-5 | NA | |
| 143 | hsa-miR-375 | SNAI2 | 0.62 | 0.1492 | -0.55 | 0.05725 | miRanda | -0.28 | 0 | NA | |
| 144 | hsa-miR-429 | SNAI2 | 2.38 | 0 | -0.55 | 0.05725 | PITA; miRanda; miRNATAP | -0.34 | 0 | NA | |
| 145 | hsa-miR-532-5p | SNAI2 | 0.58 | 0.00232 | -0.55 | 0.05725 | PITA | -0.39 | 0 | NA | |
| 146 | hsa-miR-96-5p | SNAI2 | 3.04 | 0 | -0.55 | 0.05725 | miRNATAP | -0.27 | 0 | NA | |
| 147 | hsa-miR-335-3p | SOX9 | 1.51 | 0 | 0.5 | 0.27822 | mirMAP | -0.27 | 0.00601 | NA | |
| 148 | hsa-miR-590-3p | SOX9 | 0.84 | 0.00129 | 0.5 | 0.27822 | PITA; miRanda; mirMAP; miRNATAP | -0.2 | 0.03621 | NA | |
| 149 | hsa-miR-107 | TGFB2 | 0.66 | 0 | -1.55 | 1.0E-5 | miRanda | -0.36 | 0.00389 | NA | |
| 150 | hsa-miR-130b-3p | TGFB2 | 1.83 | 0 | -1.55 | 1.0E-5 | MirTarget; miRNATAP | -0.15 | 0.02513 | NA |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | EPITHELIAL TO MESENCHYMAL TRANSITION | 28 | 56 | 8.853e-76 | 4.119e-72 |
| 2 | MESENCHYMAL CELL DIFFERENTIATION | 28 | 134 | 6.6e-63 | 1.536e-59 |
| 3 | MESENCHYME DEVELOPMENT | 28 | 190 | 2.989e-58 | 3.476e-55 |
| 4 | STEM CELL DIFFERENTIATION | 28 | 190 | 2.989e-58 | 3.476e-55 |
| 5 | CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 28 | 513 | 1.37e-45 | 1.275e-42 |
| 6 | CELLULAR COMPONENT MORPHOGENESIS | 28 | 900 | 1.3e-38 | 1.008e-35 |
| 7 | CELL DEVELOPMENT | 28 | 1426 | 6.01e-33 | 3.995e-30 |
| 8 | TISSUE DEVELOPMENT | 28 | 1518 | 3.517e-32 | 2.046e-29 |
| 9 | TISSUE MORPHOGENESIS | 20 | 533 | 5.859e-26 | 3.029e-23 |
| 10 | MESENCHYME MORPHOGENESIS | 11 | 38 | 4.945e-24 | 2.301e-21 |
| 11 | MORPHOGENESIS OF AN EPITHELIUM | 17 | 400 | 1.642e-22 | 6.946e-20 |
| 12 | EMBRYONIC ORGAN DEVELOPMENT | 17 | 406 | 2.119e-22 | 8.217e-20 |
| 13 | RESPONSE TO GROWTH FACTOR | 17 | 475 | 3.096e-21 | 1.108e-18 |
| 14 | ANATOMICAL STRUCTURE FORMATION INVOLVED IN MORPHOGENESIS | 20 | 957 | 7.059e-21 | 2.346e-18 |
| 15 | GLAND DEVELOPMENT | 16 | 395 | 9.712e-21 | 3.013e-18 |
| 16 | REGULATION OF STEM CELL DIFFERENTIATION | 12 | 113 | 1.637e-20 | 4.76e-18 |
| 17 | REGULATION OF ANATOMICAL STRUCTURE MORPHOGENESIS | 20 | 1021 | 2.543e-20 | 6.961e-18 |
| 18 | ORGAN MORPHOGENESIS | 19 | 841 | 2.817e-20 | 7.282e-18 |
| 19 | REGULATION OF OSSIFICATION | 13 | 178 | 4.7e-20 | 1.151e-17 |
| 20 | EMBRYO DEVELOPMENT | 19 | 894 | 8.895e-20 | 2.069e-17 |
| 21 | POSITIVE REGULATION OF DEVELOPMENTAL PROCESS | 20 | 1142 | 2.321e-19 | 5.144e-17 |
| 22 | HEART MORPHOGENESIS | 13 | 212 | 4.792e-19 | 1.013e-16 |
| 23 | REGULATION OF CARTILAGE DEVELOPMENT | 10 | 63 | 5.704e-19 | 1.154e-16 |
| 24 | POSITIVE REGULATION OF GENE EXPRESSION | 22 | 1733 | 8.535e-19 | 1.655e-16 |
| 25 | NEGATIVE REGULATION OF GENE EXPRESSION | 21 | 1493 | 1.345e-18 | 2.406e-16 |
| 26 | REGULATION OF CELL DIFFERENTIATION | 21 | 1492 | 1.326e-18 | 2.406e-16 |
| 27 | EMBRYONIC MORPHOGENESIS | 16 | 539 | 1.403e-18 | 2.418e-16 |
| 28 | REGULATION OF ORGAN MORPHOGENESIS | 13 | 242 | 2.749e-18 | 4.568e-16 |
| 29 | CARDIAC EPITHELIAL TO MESENCHYMAL TRANSITION | 8 | 24 | 3.554e-18 | 5.703e-16 |
| 30 | EPITHELIUM DEVELOPMENT | 18 | 945 | 9.879e-18 | 1.532e-15 |
| 31 | POSITIVE REGULATION OF STEM CELL DIFFERENTIATION | 9 | 50 | 1.186e-17 | 1.781e-15 |
| 32 | REGULATION OF EPITHELIAL CELL PROLIFERATION | 13 | 285 | 2.351e-17 | 3.418e-15 |
| 33 | POSITIVE REGULATION OF CELL DIFFERENTIATION | 17 | 823 | 3.322e-17 | 4.684e-15 |
| 34 | REGULATION OF CELL DEVELOPMENT | 17 | 836 | 4.317e-17 | 5.909e-15 |
| 35 | REGULATION OF CELL PROLIFERATION | 20 | 1496 | 4.633e-17 | 6.16e-15 |
| 36 | NEGATIVE REGULATION OF NITROGEN COMPOUND METABOLIC PROCESS | 20 | 1517 | 6.082e-17 | 7.86e-15 |
| 37 | POSITIVE REGULATION OF BIOSYNTHETIC PROCESS | 21 | 1805 | 6.621e-17 | 8.326e-15 |
| 38 | TUBE DEVELOPMENT | 15 | 552 | 9.161e-17 | 1.122e-14 |
| 39 | EMBRYO DEVELOPMENT ENDING IN BIRTH OR EGG HATCHING | 15 | 554 | 9.666e-17 | 1.153e-14 |
| 40 | TUBE MORPHOGENESIS | 13 | 323 | 1.204e-16 | 1.4e-14 |
| 41 | MESODERM MORPHOGENESIS | 9 | 66 | 1.729e-16 | 1.962e-14 |
| 42 | LOCOMOTION | 18 | 1114 | 1.8e-16 | 1.994e-14 |
| 43 | FORMATION OF PRIMARY GERM LAYER | 10 | 110 | 2.014e-16 | 2.179e-14 |
| 44 | SKELETAL SYSTEM DEVELOPMENT | 14 | 455 | 2.448e-16 | 2.589e-14 |
| 45 | POSITIVE REGULATION OF CELLULAR COMPONENT ORGANIZATION | 18 | 1152 | 3.244e-16 | 3.355e-14 |
| 46 | HEART DEVELOPMENT | 14 | 466 | 3.412e-16 | 3.451e-14 |
| 47 | REGULATION OF MULTICELLULAR ORGANISMAL DEVELOPMENT | 20 | 1672 | 4.04e-16 | 3.916e-14 |
| 48 | REGULATION OF CELLULAR COMPONENT MOVEMENT | 16 | 771 | 4.032e-16 | 3.916e-14 |
| 49 | CARDIOVASCULAR SYSTEM DEVELOPMENT | 16 | 788 | 5.679e-16 | 5.284e-14 |
| 50 | CIRCULATORY SYSTEM DEVELOPMENT | 16 | 788 | 5.679e-16 | 5.284e-14 |
| 51 | NEGATIVE REGULATION OF MULTICELLULAR ORGANISMAL PROCESS | 17 | 983 | 6.415e-16 | 5.853e-14 |
| 52 | NEGATIVE REGULATION OF DEVELOPMENTAL PROCESS | 16 | 801 | 7.34e-16 | 6.543e-14 |
| 53 | SENSORY ORGAN DEVELOPMENT | 14 | 493 | 7.453e-16 | 6.543e-14 |
| 54 | ENDOCARDIAL CUSHION MORPHOGENESIS | 7 | 22 | 7.85e-16 | 6.764e-14 |
| 55 | CELLULAR RESPONSE TO ENDOGENOUS STIMULUS | 17 | 1008 | 9.733e-16 | 8.234e-14 |
| 56 | CONNECTIVE TISSUE DEVELOPMENT | 11 | 194 | 1e-15 | 8.309e-14 |
| 57 | CELL MOTILITY | 16 | 835 | 1.408e-15 | 1.13e-13 |
| 58 | LOCALIZATION OF CELL | 16 | 835 | 1.408e-15 | 1.13e-13 |
| 59 | MOVEMENT OF CELL OR SUBCELLULAR COMPONENT | 18 | 1275 | 1.918e-15 | 1.513e-13 |
| 60 | REPRODUCTIVE SYSTEM DEVELOPMENT | 13 | 408 | 2.487e-15 | 1.929e-13 |
| 61 | REGULATION OF STEM CELL PROLIFERATION | 9 | 88 | 2.619e-15 | 1.997e-13 |
| 62 | NEGATIVE REGULATION OF CELL DEATH | 16 | 872 | 2.774e-15 | 2.082e-13 |
| 63 | NEGATIVE REGULATION OF CARTILAGE DEVELOPMENT | 7 | 26 | 3.017e-15 | 2.228e-13 |
| 64 | PATTERN SPECIFICATION PROCESS | 13 | 418 | 3.399e-15 | 2.471e-13 |
| 65 | POSITIVE REGULATION OF LOCOMOTION | 13 | 420 | 3.615e-15 | 2.588e-13 |
| 66 | CARTILAGE DEVELOPMENT | 10 | 147 | 3.955e-15 | 2.747e-13 |
| 67 | IN UTERO EMBRYONIC DEVELOPMENT | 12 | 311 | 3.94e-15 | 2.747e-13 |
| 68 | REGULATION OF MORPHOGENESIS OF A BRANCHING STRUCTURE | 8 | 53 | 4.174e-15 | 2.856e-13 |
| 69 | NEGATIVE REGULATION OF RESPONSE TO STIMULUS | 18 | 1360 | 5.918e-15 | 3.99e-13 |
| 70 | GASTRULATION | 10 | 155 | 6.785e-15 | 4.51e-13 |
| 71 | REGULATION OF CANONICAL WNT SIGNALING PATHWAY | 11 | 236 | 8.8e-15 | 5.767e-13 |
| 72 | POSITIVE REGULATION OF MULTICELLULAR ORGANISMAL PROCESS | 18 | 1395 | 9.211e-15 | 5.953e-13 |
| 73 | ENDOCARDIAL CUSHION DEVELOPMENT | 7 | 32 | 1.535e-14 | 9.773e-13 |
| 74 | NEGATIVE REGULATION OF CELL COMMUNICATION | 17 | 1192 | 1.554e-14 | 9.773e-13 |
| 75 | POSITIVE REGULATION OF CELL DEVELOPMENT | 13 | 472 | 1.622e-14 | 1.007e-12 |
| 76 | REGULATION OF CELL DEATH | 18 | 1472 | 2.344e-14 | 1.435e-12 |
| 77 | POSITIVE REGULATION OF EPITHELIAL TO MESENCHYMAL TRANSITION | 7 | 34 | 2.449e-14 | 1.48e-12 |
| 78 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 16 | 1004 | 2.493e-14 | 1.487e-12 |
| 79 | POSITIVE REGULATION OF CELL PROLIFERATION | 15 | 814 | 2.803e-14 | 1.651e-12 |
| 80 | NEGATIVE REGULATION OF CELL PROLIFERATION | 14 | 643 | 2.902e-14 | 1.688e-12 |
| 81 | REGULATION OF EPITHELIAL TO MESENCHYMAL TRANSITION | 8 | 67 | 3.034e-14 | 1.743e-12 |
| 82 | REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 19 | 1784 | 3.28e-14 | 1.861e-12 |
| 83 | MESODERM DEVELOPMENT | 9 | 118 | 3.993e-14 | 2.239e-12 |
| 84 | CELL PROLIFERATION | 14 | 672 | 5.311e-14 | 2.942e-12 |
| 85 | EMBRYONIC ORGAN MORPHOGENESIS | 11 | 279 | 5.565e-14 | 3.046e-12 |
| 86 | CELLULAR RESPONSE TO ORGANIC SUBSTANCE | 19 | 1848 | 6.239e-14 | 3.376e-12 |
| 87 | REGULATION OF BINDING | 11 | 283 | 6.506e-14 | 3.48e-12 |
| 88 | UROGENITAL SYSTEM DEVELOPMENT | 11 | 299 | 1.189e-13 | 6.287e-12 |
| 89 | REGULATION OF WNT SIGNALING PATHWAY | 11 | 310 | 1.766e-13 | 9.232e-12 |
| 90 | REGIONALIZATION | 11 | 311 | 1.829e-13 | 9.456e-12 |
| 91 | PALATE DEVELOPMENT | 8 | 85 | 2.203e-13 | 1.126e-11 |
| 92 | DIGESTIVE SYSTEM DEVELOPMENT | 9 | 148 | 3.189e-13 | 1.597e-11 |
| 93 | CELL FATE COMMITMENT | 10 | 227 | 3.193e-13 | 1.597e-11 |
| 94 | DIGESTIVE TRACT MORPHOGENESIS | 7 | 48 | 3.309e-13 | 1.638e-11 |
| 95 | DEVELOPMENTAL PROCESS INVOLVED IN REPRODUCTION | 13 | 602 | 3.624e-13 | 1.775e-11 |
| 96 | RESPONSE TO ENDOGENOUS STIMULUS | 17 | 1450 | 3.846e-13 | 1.864e-11 |
| 97 | NEGATIVE REGULATION OF CELL DIFFERENTIATION | 13 | 609 | 4.197e-13 | 2.013e-11 |
| 98 | REGULATION OF CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 11 | 337 | 4.395e-13 | 2.087e-11 |
| 99 | POSITIVE REGULATION OF EPITHELIAL CELL PROLIFERATION | 9 | 154 | 4.581e-13 | 2.153e-11 |
| 100 | NEGATIVE REGULATION OF CANONICAL WNT SIGNALING PATHWAY | 9 | 162 | 7.264e-13 | 3.346e-11 |
| 101 | POSITIVE REGULATION OF CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 9 | 162 | 7.264e-13 | 3.346e-11 |
| 102 | MORPHOGENESIS OF A BRANCHING STRUCTURE | 9 | 167 | 9.574e-13 | 4.368e-11 |
| 103 | EPITHELIAL CELL DIFFERENTIATION | 12 | 495 | 9.829e-13 | 4.44e-11 |
| 104 | NEURON DIFFERENTIATION | 14 | 874 | 1.895e-12 | 8.478e-11 |
| 105 | POSITIVE REGULATION OF STEM CELL PROLIFERATION | 7 | 61 | 1.937e-12 | 8.585e-11 |
| 106 | REGULATION OF HEART MORPHOGENESIS | 6 | 29 | 1.972e-12 | 8.587e-11 |
| 107 | COLUMNAR CUBOIDAL EPITHELIAL CELL DIFFERENTIATION | 8 | 111 | 1.975e-12 | 8.587e-11 |
| 108 | REGULATION OF OSTEOBLAST DIFFERENTIATION | 8 | 112 | 2.125e-12 | 9.153e-11 |
| 109 | ANTERIOR POSTERIOR PATTERN SPECIFICATION | 9 | 194 | 3.717e-12 | 1.572e-10 |
| 110 | GROWTH | 11 | 410 | 3.693e-12 | 1.572e-10 |
| 111 | NEUROGENESIS | 16 | 1402 | 4.252e-12 | 1.758e-10 |
| 112 | RESPIRATORY SYSTEM DEVELOPMENT | 9 | 197 | 4.269e-12 | 1.758e-10 |
| 113 | NEGATIVE REGULATION OF WNT SIGNALING PATHWAY | 9 | 197 | 4.269e-12 | 1.758e-10 |
| 114 | NEGATIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 13 | 740 | 4.921e-12 | 2.008e-10 |
| 115 | KIDNEY EPITHELIUM DEVELOPMENT | 8 | 125 | 5.195e-12 | 2.102e-10 |
| 116 | SKIN EPIDERMIS DEVELOPMENT | 7 | 71 | 5.852e-12 | 2.347e-10 |
| 117 | REGULATION OF TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 9 | 207 | 6.669e-12 | 2.652e-10 |
| 118 | BRANCHING MORPHOGENESIS OF AN EPITHELIAL TUBE | 8 | 131 | 7.599e-12 | 2.996e-10 |
| 119 | SKIN DEVELOPMENT | 9 | 211 | 7.923e-12 | 3.098e-10 |
| 120 | CELL SURFACE RECEPTOR SIGNALING PATHWAY INVOLVED IN HEART DEVELOPMENT | 5 | 16 | 1.594e-11 | 6.18e-10 |
| 121 | REGULATION OF CELL ADHESION | 12 | 629 | 1.62e-11 | 6.23e-10 |
| 122 | REGULATION OF CELLULAR RESPONSE TO GROWTH FACTOR STIMULUS | 9 | 229 | 1.652e-11 | 6.302e-10 |
| 123 | REGULATION OF GROWTH | 12 | 633 | 1.744e-11 | 6.598e-10 |
| 124 | HAIR CYCLE | 7 | 83 | 1.807e-11 | 6.726e-10 |
| 125 | MOLTING CYCLE | 7 | 83 | 1.807e-11 | 6.726e-10 |
| 126 | PROSTATE GLAND DEVELOPMENT | 6 | 41 | 1.845e-11 | 6.814e-10 |
| 127 | FOREBRAIN DEVELOPMENT | 10 | 357 | 2.857e-11 | 1.047e-09 |
| 128 | MESONEPHROS DEVELOPMENT | 7 | 90 | 3.23e-11 | 1.174e-09 |
| 129 | BLOOD VESSEL MORPHOGENESIS | 10 | 364 | 3.458e-11 | 1.247e-09 |
| 130 | OSSIFICATION | 9 | 251 | 3.754e-11 | 1.344e-09 |
| 131 | POSITIVE REGULATION OF RESPONSE TO STIMULUS | 17 | 1929 | 3.85e-11 | 1.368e-09 |
| 132 | REGULATION OF INTRACELLULAR SIGNAL TRANSDUCTION | 16 | 1656 | 5.301e-11 | 1.869e-09 |
| 133 | GLAND MORPHOGENESIS | 7 | 97 | 5.519e-11 | 1.931e-09 |
| 134 | REGULATION OF PROTEIN BINDING | 8 | 168 | 5.648e-11 | 1.961e-09 |
| 135 | APPENDAGE DEVELOPMENT | 8 | 169 | 5.923e-11 | 2.026e-09 |
| 136 | LIMB DEVELOPMENT | 8 | 169 | 5.923e-11 | 2.026e-09 |
| 137 | POSITIVE REGULATION OF TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 7 | 100 | 6.858e-11 | 2.329e-09 |
| 138 | REGULATION OF MUSCLE TISSUE DEVELOPMENT | 7 | 103 | 8.465e-11 | 2.834e-09 |
| 139 | REGULATION OF MUSCLE ORGAN DEVELOPMENT | 7 | 103 | 8.465e-11 | 2.834e-09 |
| 140 | CARDIAC CHAMBER MORPHOGENESIS | 7 | 104 | 9.067e-11 | 2.971e-09 |
| 141 | DEVELOPMENTAL GROWTH INVOLVED IN MORPHOGENESIS | 7 | 104 | 9.067e-11 | 2.971e-09 |
| 142 | REGULATION OF CELL MORPHOGENESIS | 11 | 552 | 9.001e-11 | 2.971e-09 |
| 143 | CARDIAC VENTRICLE DEVELOPMENT | 7 | 106 | 1.038e-10 | 3.378e-09 |
| 144 | CARDIAC MUSCLE TISSUE MORPHOGENESIS | 6 | 54 | 1.047e-10 | 3.383e-09 |
| 145 | ANGIOGENESIS | 9 | 293 | 1.488e-10 | 4.775e-09 |
| 146 | REGULATION OF CELLULAR COMPONENT BIOGENESIS | 12 | 767 | 1.607e-10 | 5.121e-09 |
| 147 | NEGATIVE REGULATION OF EPITHELIAL CELL PROLIFERATION | 7 | 115 | 1.851e-10 | 5.781e-09 |
| 148 | MUSCLE STRUCTURE DEVELOPMENT | 10 | 432 | 1.848e-10 | 5.781e-09 |
| 149 | NEPHRON DEVELOPMENT | 7 | 115 | 1.851e-10 | 5.781e-09 |
| 150 | STEM CELL PROLIFERATION | 6 | 60 | 2.018e-10 | 6.26e-09 |
| 151 | POSITIVE REGULATION OF CELL DEATH | 11 | 605 | 2.386e-10 | 7.353e-09 |
| 152 | POSITIVE REGULATION OF CELL COMMUNICATION | 15 | 1532 | 2.472e-10 | 7.568e-09 |
| 153 | RESPONSE TO OXYGEN LEVELS | 9 | 311 | 2.524e-10 | 7.675e-09 |
| 154 | EMBRYONIC SKELETAL SYSTEM DEVELOPMENT | 7 | 122 | 2.812e-10 | 8.449e-09 |
| 155 | RESPONSE TO ABIOTIC STIMULUS | 13 | 1024 | 2.815e-10 | 8.449e-09 |
| 156 | AXIS ELONGATION | 5 | 27 | 2.915e-10 | 8.693e-09 |
| 157 | ENDOCRINE SYSTEM DEVELOPMENT | 7 | 123 | 2.979e-10 | 8.829e-09 |
| 158 | REPRODUCTION | 14 | 1297 | 3.685e-10 | 1.085e-08 |
| 159 | VASCULATURE DEVELOPMENT | 10 | 469 | 4.109e-10 | 1.203e-08 |
| 160 | REGULATION OF REPRODUCTIVE PROCESS | 7 | 129 | 4.169e-10 | 1.212e-08 |
| 161 | EPIDERMIS MORPHOGENESIS | 5 | 29 | 4.279e-10 | 1.237e-08 |
| 162 | DEVELOPMENTAL GROWTH | 9 | 333 | 4.616e-10 | 1.326e-08 |
| 163 | MUSCLE ORGAN MORPHOGENESIS | 6 | 70 | 5.235e-10 | 1.494e-08 |
| 164 | MORPHOGENESIS OF EMBRYONIC EPITHELIUM | 7 | 134 | 5.449e-10 | 1.546e-08 |
| 165 | REGULATION OF ORGAN FORMATION | 5 | 32 | 7.236e-10 | 2.04e-08 |
| 166 | CARDIAC MUSCLE TISSUE DEVELOPMENT | 7 | 140 | 7.415e-10 | 2.078e-08 |
| 167 | RESPONSE TO LIPID | 12 | 888 | 8.596e-10 | 2.395e-08 |
| 168 | CARDIAC CHAMBER DEVELOPMENT | 7 | 144 | 9.037e-10 | 2.488e-08 |
| 169 | RESPONSE TO TRANSFORMING GROWTH FACTOR BETA | 7 | 144 | 9.037e-10 | 2.488e-08 |
| 170 | SENSORY ORGAN MORPHOGENESIS | 8 | 239 | 9.355e-10 | 2.56e-08 |
| 171 | REGULATION OF CELLULAR RESPONSE TO STRESS | 11 | 691 | 9.722e-10 | 2.645e-08 |
| 172 | REGULATION OF APOPTOTIC SIGNALING PATHWAY | 9 | 363 | 9.866e-10 | 2.669e-08 |
| 173 | REGULATION OF MESENCHYMAL CELL PROLIFERATION | 5 | 34 | 9.979e-10 | 2.684e-08 |
| 174 | RENAL TUBULE DEVELOPMENT | 6 | 78 | 1.018e-09 | 2.691e-08 |
| 175 | EPITHELIAL TO MESENCHYMAL TRANSITION INVOLVED IN ENDOCARDIAL CUSHION FORMATION | 4 | 11 | 1.007e-09 | 2.691e-08 |
| 176 | SOMITE DEVELOPMENT | 6 | 78 | 1.018e-09 | 2.691e-08 |
| 177 | HEAD DEVELOPMENT | 11 | 709 | 1.274e-09 | 3.349e-08 |
| 178 | AMEBOIDAL TYPE CELL MIGRATION | 7 | 154 | 1.447e-09 | 3.782e-08 |
| 179 | EPIDERMIS DEVELOPMENT | 8 | 253 | 1.466e-09 | 3.812e-08 |
| 180 | EMBRYONIC PLACENTA DEVELOPMENT | 6 | 83 | 1.489e-09 | 3.848e-08 |
| 181 | POSITIVE REGULATION OF OSSIFICATION | 6 | 84 | 1.602e-09 | 4.117e-08 |
| 182 | PATHWAY RESTRICTED SMAD PROTEIN PHOSPHORYLATION | 4 | 13 | 2.178e-09 | 5.537e-08 |
| 183 | MESENCHYMAL CELL PROLIFERATION | 4 | 13 | 2.178e-09 | 5.537e-08 |
| 184 | MUSCLE TISSUE DEVELOPMENT | 8 | 275 | 2.827e-09 | 7.148e-08 |
| 185 | NEPHRON EPITHELIUM DEVELOPMENT | 6 | 93 | 2.978e-09 | 7.49e-08 |
| 186 | CANONICAL WNT SIGNALING PATHWAY | 6 | 95 | 3.389e-09 | 8.478e-08 |
| 187 | ENDOCARDIAL CUSHION FORMATION | 4 | 15 | 4.149e-09 | 1.027e-07 |
| 188 | REGULATION OF CELL PROLIFERATION INVOLVED IN HEART MORPHOGENESIS | 4 | 15 | 4.149e-09 | 1.027e-07 |
| 189 | POSITIVE REGULATION OF PHOSPHATE METABOLIC PROCESS | 12 | 1036 | 4.93e-09 | 1.195e-07 |
| 190 | POSITIVE REGULATION OF PHOSPHORUS METABOLIC PROCESS | 12 | 1036 | 4.93e-09 | 1.195e-07 |
| 191 | REGULATION OF CHONDROCYTE DIFFERENTIATION | 5 | 46 | 4.859e-09 | 1.195e-07 |
| 192 | CAMERA TYPE EYE MORPHOGENESIS | 6 | 101 | 4.914e-09 | 1.195e-07 |
| 193 | PARAXIAL MESODERM DEVELOPMENT | 4 | 16 | 5.527e-09 | 1.333e-07 |
| 194 | NEGATIVE REGULATION OF CELL DEVELOPMENT | 8 | 303 | 6.045e-09 | 1.446e-07 |
| 195 | REGULATION OF CARDIAC MUSCLE TISSUE DEVELOPMENT | 5 | 48 | 6.059e-09 | 1.446e-07 |
| 196 | ODONTOGENESIS | 6 | 105 | 6.217e-09 | 1.476e-07 |
| 197 | TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 7 | 190 | 6.253e-09 | 1.477e-07 |
| 198 | REGULATION OF PHOSPHORUS METABOLIC PROCESS | 14 | 1618 | 6.603e-09 | 1.552e-07 |
| 199 | EAR DEVELOPMENT | 7 | 195 | 7.486e-09 | 1.75e-07 |
| 200 | NEGATIVE REGULATION OF APOPTOTIC SIGNALING PATHWAY | 7 | 200 | 8.921e-09 | 2.076e-07 |
| 201 | NOTOCHORD DEVELOPMENT | 4 | 18 | 9.275e-09 | 2.137e-07 |
| 202 | SKELETAL SYSTEM MORPHOGENESIS | 7 | 201 | 9.235e-09 | 2.137e-07 |
| 203 | MESONEPHRIC TUBULE MORPHOGENESIS | 5 | 53 | 1.011e-08 | 2.316e-07 |
| 204 | NOTCH SIGNALING PATHWAY | 6 | 114 | 1.021e-08 | 2.33e-07 |
| 205 | EYE DEVELOPMENT | 8 | 326 | 1.07e-08 | 2.429e-07 |
| 206 | REGULATION OF MAPK CASCADE | 10 | 660 | 1.097e-08 | 2.478e-07 |
| 207 | CENTRAL NERVOUS SYSTEM DEVELOPMENT | 11 | 872 | 1.104e-08 | 2.483e-07 |
| 208 | OUTFLOW TRACT MORPHOGENESIS | 5 | 56 | 1.341e-08 | 2.997e-07 |
| 209 | REGULATION OF PROTEIN MODIFICATION PROCESS | 14 | 1710 | 1.346e-08 | 2.997e-07 |
| 210 | POSITIVE REGULATION OF PROTEIN MODIFICATION PROCESS | 12 | 1135 | 1.373e-08 | 3.042e-07 |
| 211 | NEGATIVE REGULATION OF CHONDROCYTE DIFFERENTIATION | 4 | 20 | 1.466e-08 | 3.232e-07 |
| 212 | EMBRYONIC PATTERN SPECIFICATION | 5 | 58 | 1.606e-08 | 3.524e-07 |
| 213 | ENZYME LINKED RECEPTOR PROTEIN SIGNALING PATHWAY | 10 | 689 | 1.651e-08 | 3.606e-07 |
| 214 | OSTEOBLAST DIFFERENTIATION | 6 | 126 | 1.865e-08 | 4.055e-07 |
| 215 | REGULATION OF PATHWAY RESTRICTED SMAD PROTEIN PHOSPHORYLATION | 5 | 60 | 1.91e-08 | 4.134e-07 |
| 216 | POSITIVE REGULATION OF BINDING | 6 | 127 | 1.956e-08 | 4.213e-07 |
| 217 | TUBE FORMATION | 6 | 129 | 2.148e-08 | 4.606e-07 |
| 218 | SOMATIC STEM CELL DIVISION | 4 | 22 | 2.209e-08 | 4.714e-07 |
| 219 | CARDIAC VENTRICLE MORPHOGENESIS | 5 | 62 | 2.259e-08 | 4.8e-07 |
| 220 | NEUROEPITHELIAL CELL DIFFERENTIATION | 5 | 63 | 2.451e-08 | 5.185e-07 |
| 221 | PROTEIN PHOSPHORYLATION | 11 | 944 | 2.506e-08 | 5.276e-07 |
| 222 | REGULATION OF CELL CYCLE | 11 | 949 | 2.646e-08 | 5.546e-07 |
| 223 | PROSTATE GLAND MORPHOGENESIS | 4 | 23 | 2.671e-08 | 5.574e-07 |
| 224 | MUSCLE CELL DIFFERENTIATION | 7 | 237 | 2.876e-08 | 5.975e-07 |
| 225 | POSITIVE REGULATION OF GROWTH | 7 | 238 | 2.961e-08 | 6.096e-07 |
| 226 | EYE MORPHOGENESIS | 6 | 136 | 2.948e-08 | 6.096e-07 |
| 227 | RESPONSE TO EXTERNAL STIMULUS | 14 | 1821 | 3.012e-08 | 6.174e-07 |
| 228 | PLACENTA DEVELOPMENT | 6 | 138 | 3.217e-08 | 6.565e-07 |
| 229 | ORGAN GROWTH | 5 | 68 | 3.618e-08 | 7.352e-07 |
| 230 | REGULATION OF NERVOUS SYSTEM DEVELOPMENT | 10 | 750 | 3.682e-08 | 7.45e-07 |
| 231 | REGULATION OF FIBROBLAST GROWTH FACTOR RECEPTOR SIGNALING PATHWAY | 4 | 25 | 3.809e-08 | 7.672e-07 |
| 232 | NEGATIVE REGULATION OF OSSIFICATION | 5 | 69 | 3.897e-08 | 7.816e-07 |
| 233 | ENDODERM DEVELOPMENT | 5 | 71 | 4.506e-08 | 8.961e-07 |
| 234 | NEGATIVE REGULATION OF EMBRYONIC DEVELOPMENT | 4 | 26 | 4.497e-08 | 8.961e-07 |
| 235 | EMBRYONIC HEART TUBE DEVELOPMENT | 5 | 73 | 5.189e-08 | 1.027e-06 |
| 236 | POSITIVE REGULATION OF MESENCHYMAL CELL PROLIFERATION | 4 | 27 | 5.274e-08 | 1.035e-06 |
| 237 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER INVOLVED IN CELLULAR RESPONSE TO CHEMICAL STIMULUS | 4 | 27 | 5.274e-08 | 1.035e-06 |
| 238 | NEGATIVE REGULATION OF NERVOUS SYSTEM DEVELOPMENT | 7 | 262 | 5.72e-08 | 1.115e-06 |
| 239 | REGULATION OF MUSCLE CELL DIFFERENTIATION | 6 | 152 | 5.727e-08 | 1.115e-06 |
| 240 | POSITIVE REGULATION OF CELLULAR COMPONENT BIOGENESIS | 8 | 406 | 5.863e-08 | 1.137e-06 |
| 241 | IMMUNE SYSTEM DEVELOPMENT | 9 | 582 | 5.928e-08 | 1.144e-06 |
| 242 | VENTRICULAR SEPTUM MORPHOGENESIS | 4 | 28 | 6.147e-08 | 1.182e-06 |
| 243 | DIENCEPHALON DEVELOPMENT | 5 | 77 | 6.799e-08 | 1.302e-06 |
| 244 | REGULATION OF CARDIAC MUSCLE CELL PROLIFERATION | 4 | 29 | 7.124e-08 | 1.347e-06 |
| 245 | STEM CELL DIVISION | 4 | 29 | 7.124e-08 | 1.347e-06 |
| 246 | POSITIVE REGULATION OF CARTILAGE DEVELOPMENT | 4 | 29 | 7.124e-08 | 1.347e-06 |
| 247 | KIDNEY MORPHOGENESIS | 5 | 82 | 9.344e-08 | 1.76e-06 |
| 248 | REGULATION OF EPITHELIAL CELL MIGRATION | 6 | 166 | 9.671e-08 | 1.814e-06 |
| 249 | POSITIVE REGULATION OF NERVOUS SYSTEM DEVELOPMENT | 8 | 437 | 1.032e-07 | 1.929e-06 |
| 250 | ORGAN FORMATION | 4 | 34 | 1.384e-07 | 2.566e-06 |
| 251 | HEART VALVE DEVELOPMENT | 4 | 34 | 1.384e-07 | 2.566e-06 |
| 252 | POSITIVE REGULATION OF INTRACELLULAR SIGNAL TRANSDUCTION | 10 | 876 | 1.578e-07 | 2.914e-06 |
| 253 | EMBRYONIC SKELETAL SYSTEM MORPHOGENESIS | 5 | 93 | 1.761e-07 | 3.239e-06 |
| 254 | RESPONSE TO BMP | 5 | 94 | 1.858e-07 | 3.391e-06 |
| 255 | CELLULAR RESPONSE TO BMP STIMULUS | 5 | 94 | 1.858e-07 | 3.391e-06 |
| 256 | EPITHELIAL CELL DEVELOPMENT | 6 | 186 | 1.897e-07 | 3.447e-06 |
| 257 | NEGATIVE REGULATION OF EPITHELIAL CELL DIFFERENTIATION | 4 | 37 | 1.966e-07 | 3.559e-06 |
| 258 | POSITIVE REGULATION OF MOLECULAR FUNCTION | 13 | 1791 | 2.361e-07 | 4.258e-06 |
| 259 | RESPONSE TO ORGANIC CYCLIC COMPOUND | 10 | 917 | 2.414e-07 | 4.336e-06 |
| 260 | WNT SIGNALING PATHWAY CALCIUM MODULATING PATHWAY | 4 | 39 | 2.443e-07 | 4.356e-06 |
| 261 | GLANDULAR EPITHELIAL CELL DIFFERENTIATION | 4 | 39 | 2.443e-07 | 4.356e-06 |
| 262 | RESPONSE TO STEROID HORMONE | 8 | 497 | 2.758e-07 | 4.898e-06 |
| 263 | POSITIVE REGULATION OF PROTEIN METABOLIC PROCESS | 12 | 1492 | 2.804e-07 | 4.943e-06 |
| 264 | NEGATIVE REGULATION OF TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 5 | 102 | 2.798e-07 | 4.943e-06 |
| 265 | POSITIVE REGULATION OF EPITHELIAL CELL MIGRATION | 5 | 103 | 2.938e-07 | 5.159e-06 |
| 266 | PITUITARY GLAND DEVELOPMENT | 4 | 42 | 3.315e-07 | 5.778e-06 |
| 267 | REGULATION OF HEART GROWTH | 4 | 42 | 3.315e-07 | 5.778e-06 |
| 268 | REGULATION OF PROTEIN LOCALIZATION | 10 | 950 | 3.348e-07 | 5.813e-06 |
| 269 | PHOSPHORYLATION | 11 | 1228 | 3.638e-07 | 6.292e-06 |
| 270 | EPITHELIAL CELL DIFFERENTIATION INVOLVED IN PROSTATE GLAND DEVELOPMENT | 3 | 11 | 4.024e-07 | 6.935e-06 |
| 271 | LUNG MORPHOGENESIS | 4 | 45 | 4.401e-07 | 7.5e-06 |
| 272 | VENTRICULAR CARDIAC MUSCLE TISSUE DEVELOPMENT | 4 | 45 | 4.401e-07 | 7.5e-06 |
| 273 | EXOCRINE SYSTEM DEVELOPMENT | 4 | 45 | 4.401e-07 | 7.5e-06 |
| 274 | REGULATION OF PROTEIN LOCALIZATION TO NUCLEUS | 6 | 218 | 4.827e-07 | 8.197e-06 |
| 275 | REGULATION OF EMBRYONIC DEVELOPMENT | 5 | 114 | 4.875e-07 | 8.249e-06 |
| 276 | REGULATION OF MESENCHYMAL CELL APOPTOTIC PROCESS | 3 | 12 | 5.361e-07 | 9.038e-06 |
| 277 | MAMMARY GLAND DEVELOPMENT | 5 | 117 | 5.548e-07 | 9.32e-06 |
| 278 | CELL MORPHOGENESIS INVOLVED IN NEURON DIFFERENTIATION | 7 | 368 | 5.724e-07 | 9.557e-06 |
| 279 | POSITIVE REGULATION OF PATHWAY RESTRICTED SMAD PROTEIN PHOSPHORYLATION | 4 | 48 | 5.73e-07 | 9.557e-06 |
| 280 | REGULATION OF CELLULAR PROTEIN LOCALIZATION | 8 | 552 | 6.111e-07 | 1.016e-05 |
| 281 | CARDIAC SEPTUM MORPHOGENESIS | 4 | 49 | 6.234e-07 | 1.032e-05 |
| 282 | REGULATION OF NEURON DIFFERENTIATION | 8 | 554 | 6.28e-07 | 1.033e-05 |
| 283 | TELENCEPHALON DEVELOPMENT | 6 | 228 | 6.277e-07 | 1.033e-05 |
| 284 | NEGATIVE REGULATION OF CELLULAR RESPONSE TO GROWTH FACTOR STIMULUS | 5 | 121 | 6.558e-07 | 1.074e-05 |
| 285 | POSITIVE REGULATION OF CELL ADHESION | 7 | 376 | 6.613e-07 | 1.08e-05 |
| 286 | FACE DEVELOPMENT | 4 | 50 | 6.769e-07 | 1.101e-05 |
| 287 | CARDIOBLAST DIFFERENTIATION | 3 | 13 | 6.962e-07 | 1.129e-05 |
| 288 | REGULATION OF CYTOKINE PRODUCTION | 8 | 563 | 7.092e-07 | 1.143e-05 |
| 289 | REGULATION OF CELL CELL ADHESION | 7 | 380 | 7.099e-07 | 1.143e-05 |
| 290 | REGULATION OF VASCULATURE DEVELOPMENT | 6 | 233 | 7.126e-07 | 1.143e-05 |
| 291 | ARTERY MORPHOGENESIS | 4 | 51 | 7.338e-07 | 1.173e-05 |
| 292 | HEPATICOBILIARY SYSTEM DEVELOPMENT | 5 | 128 | 8.669e-07 | 1.381e-05 |
| 293 | CELL MIGRATION INVOLVED IN GASTRULATION | 3 | 14 | 8.853e-07 | 1.396e-05 |
| 294 | EMBRYONIC SKELETAL JOINT DEVELOPMENT | 3 | 14 | 8.853e-07 | 1.396e-05 |
| 295 | CONVERGENT EXTENSION | 3 | 14 | 8.853e-07 | 1.396e-05 |
| 296 | VENTRICULAR SEPTUM DEVELOPMENT | 4 | 54 | 9.26e-07 | 1.456e-05 |
| 297 | NEGATIVE REGULATION OF BINDING | 5 | 131 | 9.723e-07 | 1.523e-05 |
| 298 | REGULATION OF KIDNEY DEVELOPMENT | 4 | 55 | 9.977e-07 | 1.558e-05 |
| 299 | MAINTENANCE OF CELL NUMBER | 5 | 132 | 1.01e-06 | 1.571e-05 |
| 300 | NEURON PROJECTION MORPHOGENESIS | 7 | 402 | 1.034e-06 | 1.605e-05 |
| 301 | POSITIVE REGULATION OF MUSCLE TISSUE DEVELOPMENT | 4 | 56 | 1.073e-06 | 1.659e-05 |
| 302 | MORPHOGENESIS OF AN EPITHELIAL FOLD | 3 | 15 | 1.106e-06 | 1.698e-05 |
| 303 | STRIATED MUSCLE CELL PROLIFERATION | 3 | 15 | 1.106e-06 | 1.698e-05 |
| 304 | RESPONSE TO OXYGEN CONTAINING COMPOUND | 11 | 1381 | 1.173e-06 | 1.795e-05 |
| 305 | REGULATION OF GLIAL CELL DIFFERENTIATION | 4 | 59 | 1.326e-06 | 2.023e-05 |
| 306 | MORPHOGENESIS OF AN ENDOTHELIUM | 3 | 16 | 1.359e-06 | 2.067e-05 |
| 307 | POSITIVE REGULATION OF OSTEOBLAST DIFFERENTIATION | 4 | 60 | 1.42e-06 | 2.152e-05 |
| 308 | EMBRYONIC DIGIT MORPHOGENESIS | 4 | 61 | 1.518e-06 | 2.293e-05 |
| 309 | SOMITOGENESIS | 4 | 62 | 1.621e-06 | 2.441e-05 |
| 310 | BRANCH ELONGATION OF AN EPITHELIUM | 3 | 17 | 1.649e-06 | 2.46e-05 |
| 311 | NEGATIVE REGULATION OF ALCOHOL BIOSYNTHETIC PROCESS | 3 | 17 | 1.649e-06 | 2.46e-05 |
| 312 | NEGATIVE REGULATION OF STEM CELL PROLIFERATION | 3 | 17 | 1.649e-06 | 2.46e-05 |
| 313 | RESPONSE TO ESTRADIOL | 5 | 146 | 1.661e-06 | 2.47e-05 |
| 314 | MUSCLE ORGAN DEVELOPMENT | 6 | 277 | 1.95e-06 | 2.889e-05 |
| 315 | PERICARDIUM DEVELOPMENT | 3 | 18 | 1.977e-06 | 2.911e-05 |
| 316 | NEPHRON TUBULE FORMATION | 3 | 18 | 1.977e-06 | 2.911e-05 |
| 317 | ACTIVATION OF PROTEIN KINASE ACTIVITY | 6 | 279 | 2.033e-06 | 2.984e-05 |
| 318 | REGULATION OF RESPONSE TO STRESS | 11 | 1468 | 2.143e-06 | 3.135e-05 |
| 319 | RESPONSE TO HORMONE | 9 | 893 | 2.173e-06 | 3.169e-05 |
| 320 | NEURON FATE COMMITMENT | 4 | 67 | 2.216e-06 | 3.223e-05 |
| 321 | REGULATION OF CELL JUNCTION ASSEMBLY | 4 | 68 | 2.353e-06 | 3.379e-05 |
| 322 | MUSCLE CELL PROLIFERATION | 3 | 19 | 2.346e-06 | 3.379e-05 |
| 323 | POSITIVE REGULATION OF CARDIAC MUSCLE CELL PROLIFERATION | 3 | 19 | 2.346e-06 | 3.379e-05 |
| 324 | POSITIVE REGULATION OF ENDOTHELIAL CELL PROLIFERATION | 4 | 68 | 2.353e-06 | 3.379e-05 |
| 325 | REGULATION OF ORGANELLE ORGANIZATION | 10 | 1178 | 2.391e-06 | 3.423e-05 |
| 326 | CELLULAR RESPONSE TO LIPID | 7 | 457 | 2.427e-06 | 3.464e-05 |
| 327 | REGULATION OF DEVELOPMENTAL GROWTH | 6 | 289 | 2.492e-06 | 3.546e-05 |
| 328 | TRACHEA DEVELOPMENT | 3 | 20 | 2.757e-06 | 3.911e-05 |
| 329 | CELL FATE SPECIFICATION | 4 | 71 | 2.799e-06 | 3.959e-05 |
| 330 | POSITIVE REGULATION OF MAPK CASCADE | 7 | 470 | 2.922e-06 | 4.12e-05 |
| 331 | POSITIVE REGULATION OF CATALYTIC ACTIVITY | 11 | 1518 | 2.975e-06 | 4.182e-05 |
| 332 | POSITIVE REGULATION OF PROTEIN BINDING | 4 | 73 | 3.129e-06 | 4.36e-05 |
| 333 | REGULATION OF ORGAN GROWTH | 4 | 73 | 3.129e-06 | 4.36e-05 |
| 334 | REGULATION OF NEURAL PRECURSOR CELL PROLIFERATION | 4 | 73 | 3.129e-06 | 4.36e-05 |
| 335 | POSITIVE REGULATION OF NEUROBLAST PROLIFERATION | 3 | 21 | 3.214e-06 | 4.45e-05 |
| 336 | LEFT RIGHT PATTERN FORMATION | 3 | 21 | 3.214e-06 | 4.45e-05 |
| 337 | REGULATION OF TRANSFERASE ACTIVITY | 9 | 946 | 3.491e-06 | 4.764e-05 |
| 338 | ARTERY DEVELOPMENT | 4 | 75 | 3.488e-06 | 4.764e-05 |
| 339 | REGULATION OF BIOMINERAL TISSUE DEVELOPMENT | 4 | 75 | 3.488e-06 | 4.764e-05 |
| 340 | ODONTOGENESIS OF DENTIN CONTAINING TOOTH | 4 | 75 | 3.488e-06 | 4.764e-05 |
| 341 | NEURAL CREST CELL DIFFERENTIATION | 4 | 75 | 3.488e-06 | 4.764e-05 |
| 342 | EMBRYONIC PLACENTA MORPHOGENESIS | 3 | 22 | 3.718e-06 | 5.058e-05 |
| 343 | REGULATION OF BMP SIGNALING PATHWAY | 4 | 77 | 3.876e-06 | 5.258e-05 |
| 344 | GLIOGENESIS | 5 | 175 | 4.044e-06 | 5.47e-05 |
| 345 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER IN RESPONSE TO STRESS | 3 | 23 | 4.271e-06 | 5.76e-05 |
| 346 | NEGATIVE REGULATION OF PROTEIN BINDING | 4 | 79 | 4.295e-06 | 5.776e-05 |
| 347 | RESPONSE TO ACID CHEMICAL | 6 | 319 | 4.402e-06 | 5.886e-05 |
| 348 | REGULATION OF MAP KINASE ACTIVITY | 6 | 319 | 4.402e-06 | 5.886e-05 |
| 349 | CELLULAR RESPONSE TO MECHANICAL STIMULUS | 4 | 80 | 4.517e-06 | 6.022e-05 |
| 350 | TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 8 | 724 | 4.634e-06 | 6.16e-05 |
| 351 | NEGATIVE REGULATION OF STEROID METABOLIC PROCESS | 3 | 24 | 4.877e-06 | 6.446e-05 |
| 352 | EMBRYONIC CAMERA TYPE EYE MORPHOGENESIS | 3 | 24 | 4.877e-06 | 6.446e-05 |
| 353 | REGULATION OF CELLULAR LOCALIZATION | 10 | 1277 | 4.936e-06 | 6.506e-05 |
| 354 | REGULATION OF PROTEIN IMPORT | 5 | 183 | 5.031e-06 | 6.612e-05 |
| 355 | ORGAN REGENERATION | 4 | 83 | 5.233e-06 | 6.859e-05 |
| 356 | CELL JUNCTION ORGANIZATION | 5 | 185 | 5.305e-06 | 6.933e-05 |
| 357 | EPITHELIAL TUBE BRANCHING INVOLVED IN LUNG MORPHOGENESIS | 3 | 25 | 5.537e-06 | 7.136e-05 |
| 358 | LUNG CELL DIFFERENTIATION | 3 | 25 | 5.537e-06 | 7.136e-05 |
| 359 | REGULATION OF TRANSFORMING GROWTH FACTOR BETA PRODUCTION | 3 | 25 | 5.537e-06 | 7.136e-05 |
| 360 | POSITIVE REGULATION OF PEPTIDYL THREONINE PHOSPHORYLATION | 3 | 25 | 5.537e-06 | 7.136e-05 |
| 361 | POSITIVE REGULATION OF CELL CYCLE | 6 | 332 | 5.536e-06 | 7.136e-05 |
| 362 | RESPONSE TO METAL ION | 6 | 333 | 5.632e-06 | 7.239e-05 |
| 363 | CARDIAC SEPTUM DEVELOPMENT | 4 | 85 | 5.755e-06 | 7.377e-05 |
| 364 | NEGATIVE REGULATION OF NEURON DIFFERENTIATION | 5 | 191 | 6.197e-06 | 7.922e-05 |
| 365 | HEART TRABECULA MORPHOGENESIS | 3 | 26 | 6.253e-06 | 7.928e-05 |
| 366 | NEGATIVE REGULATION OF GLIAL CELL DIFFERENTIATION | 3 | 26 | 6.253e-06 | 7.928e-05 |
| 367 | HEART GROWTH | 3 | 26 | 6.253e-06 | 7.928e-05 |
| 368 | TISSUE REMODELING | 4 | 87 | 6.314e-06 | 7.984e-05 |
| 369 | DEVELOPMENTAL MATURATION | 5 | 193 | 6.52e-06 | 8.221e-05 |
| 370 | SEGMENTATION | 4 | 89 | 6.913e-06 | 8.67e-05 |
| 371 | EPITHELIAL CELL PROLIFERATION | 4 | 89 | 6.913e-06 | 8.67e-05 |
| 372 | POSITIVE REGULATION OF HEART GROWTH | 3 | 27 | 7.028e-06 | 8.72e-05 |
| 373 | DEVELOPMENTAL INDUCTION | 3 | 27 | 7.028e-06 | 8.72e-05 |
| 374 | REGULATION OF ASTROCYTE DIFFERENTIATION | 3 | 27 | 7.028e-06 | 8.72e-05 |
| 375 | RESPONSE TO LITHIUM ION | 3 | 27 | 7.028e-06 | 8.72e-05 |
| 376 | CELL CELL SIGNALING | 8 | 767 | 7.085e-06 | 8.767e-05 |
| 377 | AXIS SPECIFICATION | 4 | 90 | 7.227e-06 | 8.897e-05 |
| 378 | REGULATION OF GLIOGENESIS | 4 | 90 | 7.227e-06 | 8.897e-05 |
| 379 | WNT SIGNALING PATHWAY | 6 | 351 | 7.611e-06 | 9.344e-05 |
| 380 | REGULATION OF KINASE ACTIVITY | 8 | 776 | 7.717e-06 | 9.437e-05 |
| 381 | NEURON PROJECTION DEVELOPMENT | 7 | 545 | 7.728e-06 | 9.437e-05 |
| 382 | REGULATION OF NEUROBLAST PROLIFERATION | 3 | 28 | 7.864e-06 | 9.554e-05 |
| 383 | DOPAMINERGIC NEURON DIFFERENTIATION | 3 | 28 | 7.864e-06 | 9.554e-05 |
| 384 | REGULATION OF EXTRACELLULAR MATRIX ORGANIZATION | 3 | 29 | 8.763e-06 | 0.0001059 |
| 385 | POSITIVE REGULATION OF CARDIAC MUSCLE TISSUE DEVELOPMENT | 3 | 29 | 8.763e-06 | 0.0001059 |
| 386 | REGULATION OF CELL CYCLE PROCESS | 7 | 558 | 9.012e-06 | 0.0001086 |
| 387 | RESPONSE TO ALCOHOL | 6 | 362 | 9.075e-06 | 0.0001091 |
| 388 | CARDIOCYTE DIFFERENTIATION | 4 | 96 | 9.342e-06 | 0.000112 |
| 389 | SMOOTH MUSCLE CELL DIFFERENTIATION | 3 | 30 | 9.727e-06 | 0.0001164 |
| 390 | REGULATION OF ENDOTHELIAL CELL PROLIFERATION | 4 | 98 | 1.014e-05 | 0.000121 |
| 391 | NEGATIVE REGULATION OF MOLECULAR FUNCTION | 9 | 1079 | 1.018e-05 | 0.0001212 |
| 392 | POSITIVE REGULATION OF ORGANELLE ORGANIZATION | 7 | 573 | 1.071e-05 | 0.0001271 |
| 393 | REGULATION OF SMOOTH MUSCLE CELL PROLIFERATION | 4 | 100 | 1.099e-05 | 0.0001301 |
| 394 | RESPONSE TO ESTROGEN | 5 | 218 | 1.177e-05 | 0.000139 |
| 395 | SALIVARY GLAND DEVELOPMENT | 3 | 32 | 1.186e-05 | 0.0001394 |
| 396 | REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER IN RESPONSE TO HYPOXIA | 3 | 32 | 1.186e-05 | 0.0001394 |
| 397 | REGULATION OF NUCLEOCYTOPLASMIC TRANSPORT | 5 | 220 | 1.23e-05 | 0.0001442 |
| 398 | EMBRYONIC AXIS SPECIFICATION | 3 | 33 | 1.304e-05 | 0.0001513 |
| 399 | REGULATION OF T CELL APOPTOTIC PROCESS | 3 | 33 | 1.304e-05 | 0.0001513 |
| 400 | EMBRYONIC DIGESTIVE TRACT DEVELOPMENT | 3 | 33 | 1.304e-05 | 0.0001513 |
| 401 | EMBRYONIC EYE MORPHOGENESIS | 3 | 33 | 1.304e-05 | 0.0001513 |
| 402 | NEGATIVE REGULATION OF CELL ADHESION | 5 | 223 | 1.314e-05 | 0.0001521 |
| 403 | REGULATION OF CELL GROWTH | 6 | 391 | 1.406e-05 | 0.0001624 |
| 404 | HAIR CELL DIFFERENTIATION | 3 | 35 | 1.561e-05 | 0.0001793 |
| 405 | EMBRYONIC CAMERA TYPE EYE DEVELOPMENT | 3 | 35 | 1.561e-05 | 0.0001793 |
| 406 | PROTEIN LOCALIZATION | 11 | 1805 | 1.582e-05 | 0.0001813 |
| 407 | NEGATIVE REGULATION OF MUSCLE ORGAN DEVELOPMENT | 3 | 36 | 1.701e-05 | 0.0001935 |
| 408 | NEGATIVE REGULATION OF MUSCLE TISSUE DEVELOPMENT | 3 | 36 | 1.701e-05 | 0.0001935 |
| 409 | HEAD MORPHOGENESIS | 3 | 36 | 1.701e-05 | 0.0001935 |
| 410 | POSITIVE REGULATION OF TRANSFERASE ACTIVITY | 7 | 616 | 1.712e-05 | 0.0001943 |
| 411 | EAR MORPHOGENESIS | 4 | 112 | 1.72e-05 | 0.0001947 |
| 412 | NEGATIVE REGULATION OF GROWTH | 5 | 236 | 1.727e-05 | 0.000195 |
| 413 | NEGATIVE REGULATION OF GLIOGENESIS | 3 | 37 | 1.849e-05 | 0.0002074 |
| 414 | MYOBLAST DIFFERENTIATION | 3 | 37 | 1.849e-05 | 0.0002074 |
| 415 | REGULATION OF PEPTIDYL THREONINE PHOSPHORYLATION | 3 | 37 | 1.849e-05 | 0.0002074 |
| 416 | RESPONSE TO FIBROBLAST GROWTH FACTOR | 4 | 116 | 1.975e-05 | 0.0002209 |
| 417 | COLLAGEN FIBRIL ORGANIZATION | 3 | 38 | 2.006e-05 | 0.0002228 |
| 418 | BONE MINERALIZATION | 3 | 38 | 2.006e-05 | 0.0002228 |
| 419 | POSITIVE REGULATION OF ORGAN GROWTH | 3 | 38 | 2.006e-05 | 0.0002228 |
| 420 | CELL PART MORPHOGENESIS | 7 | 633 | 2.04e-05 | 0.0002258 |
| 421 | NEGATIVE REGULATION OF CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 4 | 117 | 2.043e-05 | 0.0002258 |
| 422 | ANATOMICAL STRUCTURE MATURATION | 3 | 39 | 2.171e-05 | 0.0002388 |
| 423 | TRABECULA MORPHOGENESIS | 3 | 39 | 2.171e-05 | 0.0002388 |
| 424 | CYTOKINE PRODUCTION | 4 | 120 | 2.258e-05 | 0.0002477 |
| 425 | POSITIVE REGULATION OF CELL MATRIX ADHESION | 3 | 40 | 2.345e-05 | 0.0002567 |
| 426 | REGULATION OF EPITHELIAL CELL DIFFERENTIATION | 4 | 122 | 2.409e-05 | 0.0002631 |
| 427 | NEGATIVE REGULATION OF CELL CYCLE | 6 | 433 | 2.501e-05 | 0.0002726 |
| 428 | NEGATIVE REGULATION OF INTRACELLULAR SIGNAL TRANSDUCTION | 6 | 437 | 2.634e-05 | 0.0002863 |
| 429 | NEGATIVE REGULATION OF BMP SIGNALING PATHWAY | 3 | 42 | 2.72e-05 | 0.0002936 |
| 430 | POSITIVE REGULATION OF NEURAL PRECURSOR CELL PROLIFERATION | 3 | 42 | 2.72e-05 | 0.0002936 |
| 431 | GENITALIA DEVELOPMENT | 3 | 42 | 2.72e-05 | 0.0002936 |
| 432 | NEGATIVE REGULATION OF LOCOMOTION | 5 | 263 | 2.904e-05 | 0.0003121 |
| 433 | CELLULAR RESPONSE TO ABIOTIC STIMULUS | 5 | 263 | 2.904e-05 | 0.0003121 |
| 434 | NEGATIVE REGULATION OF STEM CELL DIFFERENTIATION | 3 | 43 | 2.921e-05 | 0.0003132 |
| 435 | SEX DIFFERENTIATION | 5 | 266 | 3.066e-05 | 0.000328 |
| 436 | LABYRINTHINE LAYER DEVELOPMENT | 3 | 44 | 3.132e-05 | 0.0003319 |
| 437 | BODY MORPHOGENESIS | 3 | 44 | 3.132e-05 | 0.0003319 |
| 438 | NEGATIVE REGULATION OF LIPID BIOSYNTHETIC PROCESS | 3 | 44 | 3.132e-05 | 0.0003319 |
| 439 | BRANCHING INVOLVED IN URETERIC BUD MORPHOGENESIS | 3 | 44 | 3.132e-05 | 0.0003319 |
| 440 | SPROUTING ANGIOGENESIS | 3 | 45 | 3.352e-05 | 0.0003537 |
| 441 | REGULATION OF ALCOHOL BIOSYNTHETIC PROCESS | 3 | 45 | 3.352e-05 | 0.0003537 |
| 442 | NEURON DEVELOPMENT | 7 | 687 | 3.449e-05 | 0.0003631 |
| 443 | CELL DIVISION | 6 | 460 | 3.511e-05 | 0.0003688 |
| 444 | CELL GROWTH | 4 | 135 | 3.585e-05 | 0.0003748 |
| 445 | EMBRYONIC CRANIAL SKELETON MORPHOGENESIS | 3 | 46 | 3.583e-05 | 0.0003748 |
| 446 | TAXIS | 6 | 464 | 3.686e-05 | 0.0003841 |
| 447 | GLIAL CELL DIFFERENTIATION | 4 | 136 | 3.69e-05 | 0.0003841 |
| 448 | CELLULAR RESPONSE TO ORGANIC CYCLIC COMPOUND | 6 | 465 | 3.73e-05 | 0.000387 |
| 449 | PHOSPHATE CONTAINING COMPOUND METABOLIC PROCESS | 11 | 1977 | 3.735e-05 | 0.000387 |
| 450 | POSITIVE REGULATION OF NEURON APOPTOTIC PROCESS | 3 | 47 | 3.824e-05 | 0.0003954 |
| 451 | IMMUNE SYSTEM PROCESS | 11 | 1984 | 3.86e-05 | 0.0003982 |
| 452 | REGULATION OF PHOSPHATIDYLINOSITOL 3 KINASE SIGNALING | 4 | 138 | 3.907e-05 | 0.0004013 |
| 453 | NEGATIVE REGULATION OF CELL CELL ADHESION | 4 | 138 | 3.907e-05 | 0.0004013 |
| 454 | WOUND HEALING | 6 | 470 | 3.96e-05 | 0.000405 |
| 455 | REGULATION OF PROTEIN SERINE THREONINE KINASE ACTIVITY | 6 | 470 | 3.96e-05 | 0.000405 |
| 456 | ANTERIOR POSTERIOR AXIS SPECIFICATION | 3 | 48 | 4.075e-05 | 0.0004158 |
| 457 | NON CANONICAL WNT SIGNALING PATHWAY | 4 | 140 | 4.133e-05 | 0.0004208 |
| 458 | REGULATION OF STEROID BIOSYNTHETIC PROCESS | 3 | 49 | 4.336e-05 | 0.0004405 |
| 459 | EPIDERMAL CELL DIFFERENTIATION | 4 | 142 | 4.368e-05 | 0.0004428 |
| 460 | RESPONSE TO INORGANIC SUBSTANCE | 6 | 479 | 4.402e-05 | 0.0004453 |
| 461 | CELLULAR RESPONSE TO OXYGEN LEVELS | 4 | 143 | 4.49e-05 | 0.0004532 |
| 462 | POSITIVE REGULATION OF KINASE ACTIVITY | 6 | 482 | 4.558e-05 | 0.000459 |
| 463 | ENDODERM FORMATION | 3 | 50 | 4.609e-05 | 0.0004632 |
| 464 | REGULATION OF RESPONSE TO DNA DAMAGE STIMULUS | 4 | 145 | 4.74e-05 | 0.0004753 |
| 465 | POSITIVE REGULATION OF CELL GROWTH | 4 | 148 | 5.134e-05 | 0.0005126 |
| 466 | MALE SEX DIFFERENTIATION | 4 | 148 | 5.134e-05 | 0.0005126 |
| 467 | POSITIVE REGULATION OF REPRODUCTIVE PROCESS | 3 | 52 | 5.187e-05 | 0.0005157 |
| 468 | NEGATIVE REGULATION OF RESPONSE TO DNA DAMAGE STIMULUS | 3 | 52 | 5.187e-05 | 0.0005157 |
| 469 | NEURAL TUBE DEVELOPMENT | 4 | 149 | 5.271e-05 | 0.0005229 |
| 470 | REGULATION OF HYDROLASE ACTIVITY | 9 | 1327 | 5.285e-05 | 0.0005232 |
| 471 | MAMMARY GLAND EPITHELIUM DEVELOPMENT | 3 | 53 | 5.493e-05 | 0.0005427 |
| 472 | POSITIVE REGULATION OF WNT SIGNALING PATHWAY | 4 | 152 | 5.696e-05 | 0.0005608 |
| 473 | POSITIVE REGULATION OF CELL PROJECTION ORGANIZATION | 5 | 303 | 5.701e-05 | 0.0005608 |
| 474 | NEGATIVE REGULATION OF REPRODUCTIVE PROCESS | 3 | 54 | 5.811e-05 | 0.000568 |
| 475 | REGULATION OF LYMPHOCYTE APOPTOTIC PROCESS | 3 | 54 | 5.811e-05 | 0.000568 |
| 476 | EXTRACELLULAR STRUCTURE ORGANIZATION | 5 | 304 | 5.791e-05 | 0.000568 |
| 477 | REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY | 4 | 153 | 5.843e-05 | 0.0005688 |
| 478 | PALLIUM DEVELOPMENT | 4 | 153 | 5.843e-05 | 0.0005688 |
| 479 | POSITIVE REGULATION OF NEURON DIFFERENTIATION | 5 | 306 | 5.974e-05 | 0.0005803 |
| 480 | REGULATION OF PROTEIN TARGETING | 5 | 307 | 6.067e-05 | 0.0005881 |
| 481 | NEGATIVE REGULATION OF MUSCLE CELL DIFFERENTIATION | 3 | 55 | 6.14e-05 | 0.0005927 |
| 482 | CRANIAL SKELETAL SYSTEM DEVELOPMENT | 3 | 55 | 6.14e-05 | 0.0005927 |
| 483 | POSITIVE REGULATION OF DEVELOPMENTAL GROWTH | 4 | 156 | 6.302e-05 | 0.0006046 |
| 484 | CELLULAR RESPONSE TO INORGANIC SUBSTANCE | 4 | 156 | 6.302e-05 | 0.0006046 |
| 485 | PROTEIN LOCALIZATION TO NUCLEUS | 4 | 156 | 6.302e-05 | 0.0006046 |
| 486 | SMAD PROTEIN SIGNAL TRANSDUCTION | 3 | 56 | 6.482e-05 | 0.0006206 |
| 487 | POSITIVE REGULATION OF ESTABLISHMENT OF PROTEIN LOCALIZATION | 6 | 514 | 6.514e-05 | 0.0006224 |
| 488 | POSITIVE REGULATION OF EPITHELIAL CELL DIFFERENTIATION | 3 | 57 | 6.835e-05 | 0.0006517 |
| 489 | MULTICELLULAR ORGANISM REPRODUCTION | 7 | 768 | 6.998e-05 | 0.0006659 |
| 490 | REGENERATION | 4 | 161 | 7.124e-05 | 0.0006765 |
| 491 | POSITIVE REGULATION OF DNA BIOSYNTHETIC PROCESS | 3 | 59 | 7.58e-05 | 0.0007183 |
| 492 | POSITIVE REGULATION OF SMOOTH MUSCLE CELL PROLIFERATION | 3 | 60 | 7.972e-05 | 0.0007524 |
| 493 | CHONDROCYTE DIFFERENTIATION | 3 | 60 | 7.972e-05 | 0.0007524 |
| 494 | REGULATION OF VIRAL TRANSCRIPTION | 3 | 61 | 8.376e-05 | 0.0007889 |
| 495 | CHROMATIN MODIFICATION | 6 | 539 | 8.469e-05 | 0.0007961 |
| 496 | POSITIVE REGULATION OF PHOSPHATIDYLINOSITOL 3 KINASE SIGNALING | 3 | 62 | 8.794e-05 | 0.0008233 |
| 497 | EMBRYONIC HEART TUBE MORPHOGENESIS | 3 | 62 | 8.794e-05 | 0.0008233 |
| 498 | REGULATION OF CELL SUBSTRATE ADHESION | 4 | 173 | 9.413e-05 | 0.0008795 |
| 499 | CELLULAR RESPONSE TO ACID CHEMICAL | 4 | 175 | 9.841e-05 | 0.0009158 |
| 500 | REGULATION OF DNA METABOLIC PROCESS | 5 | 340 | 9.823e-05 | 0.0009158 |
| 501 | REGULATION OF TRANSPORT | 10 | 1804 | 9.984e-05 | 0.0009273 |
| 502 | PROTEIN LOCALIZATION TO ORGANELLE | 6 | 556 | 0.0001005 | 0.0009313 |
| 503 | CELLULAR RESPONSE TO RETINOIC ACID | 3 | 65 | 0.0001013 | 0.0009332 |
| 504 | REGULATION OF CHEMOKINE PRODUCTION | 3 | 65 | 0.0001013 | 0.0009332 |
| 505 | REGULATION OF RESPONSE TO OXIDATIVE STRESS | 3 | 65 | 0.0001013 | 0.0009332 |
| 506 | ENDOCARDIUM DEVELOPMENT | 2 | 11 | 0.0001031 | 0.0009374 |
| 507 | REGULATION OF EXTRACELLULAR MATRIX ASSEMBLY | 2 | 11 | 0.0001031 | 0.0009374 |
| 508 | RESPONSE TO FOLLICLE STIMULATING HORMONE | 2 | 11 | 0.0001031 | 0.0009374 |
| 509 | PROSTATE GLAND GROWTH | 2 | 11 | 0.0001031 | 0.0009374 |
| 510 | VENTRICULAR TRABECULA MYOCARDIUM MORPHOGENESIS | 2 | 11 | 0.0001031 | 0.0009374 |
| 511 | PROSTATE GLANDULAR ACINUS DEVELOPMENT | 2 | 11 | 0.0001031 | 0.0009374 |
| 512 | ENDOTHELIAL TUBE MORPHOGENESIS | 2 | 11 | 0.0001031 | 0.0009374 |
| 513 | SOMATIC STEM CELL POPULATION MAINTENANCE | 3 | 66 | 0.000106 | 0.0009577 |
| 514 | FOREBRAIN GENERATION OF NEURONS | 3 | 66 | 0.000106 | 0.0009577 |
| 515 | LENS DEVELOPMENT IN CAMERA TYPE EYE | 3 | 66 | 0.000106 | 0.0009577 |
| 516 | RESPONSE TO WOUNDING | 6 | 563 | 0.0001076 | 0.0009705 |
| 517 | POSITIVE REGULATION OF NEURON DEATH | 3 | 67 | 0.0001109 | 0.0009959 |
| 518 | REGULATION OF DNA TEMPLATED TRANSCRIPTION IN RESPONSE TO STRESS | 3 | 67 | 0.0001109 | 0.0009959 |
| 519 | RESPONSE TO ACTIVITY | 3 | 69 | 0.000121 | 0.001085 |
| 520 | CARDIAC LEFT VENTRICLE MORPHOGENESIS | 2 | 12 | 0.0001237 | 0.001088 |
| 521 | REGULATION OF TIMING OF CELL DIFFERENTIATION | 2 | 12 | 0.0001237 | 0.001088 |
| 522 | POSITIVE REGULATION OF DNA METABOLIC PROCESS | 4 | 185 | 0.000122 | 0.001088 |
| 523 | CARTILAGE MORPHOGENESIS | 2 | 12 | 0.0001237 | 0.001088 |
| 524 | REGULATION OF VITAMIN METABOLIC PROCESS | 2 | 12 | 0.0001237 | 0.001088 |
| 525 | REGULATION OF THYMOCYTE APOPTOTIC PROCESS | 2 | 12 | 0.0001237 | 0.001088 |
| 526 | REGULATION OF MACROPHAGE CYTOKINE PRODUCTION | 2 | 12 | 0.0001237 | 0.001088 |
| 527 | REGULATION OF DEVELOPMENT HETEROCHRONIC | 2 | 12 | 0.0001237 | 0.001088 |
| 528 | POSITIVE REGULATION OF ASTROCYTE DIFFERENTIATION | 2 | 12 | 0.0001237 | 0.001088 |
| 529 | NEGATIVE REGULATION OF RECEPTOR BINDING | 2 | 12 | 0.0001237 | 0.001088 |
| 530 | NEGATIVE REGULATION OF NUCLEOCYTOPLASMIC TRANSPORT | 3 | 71 | 0.0001318 | 0.001157 |
| 531 | MYELOID CELL DIFFERENTIATION | 4 | 189 | 0.0001324 | 0.001161 |
| 532 | ENDOTHELIAL CELL DIFFERENTIATION | 3 | 72 | 0.0001374 | 0.001202 |
| 533 | REGULATION OF NEURON APOPTOTIC PROCESS | 4 | 192 | 0.0001407 | 0.001228 |
| 534 | NEURONAL STEM CELL DIVISION | 2 | 13 | 0.000146 | 0.001247 |
| 535 | NEGATIVE REGULATION OF OLIGODENDROCYTE DIFFERENTIATION | 2 | 13 | 0.000146 | 0.001247 |
| 536 | PANCREAS DEVELOPMENT | 3 | 73 | 0.0001431 | 0.001247 |
| 537 | LEFT RIGHT AXIS SPECIFICATION | 2 | 13 | 0.000146 | 0.001247 |
| 538 | CELLULAR RESPONSE TO HEPATOCYTE GROWTH FACTOR STIMULUS | 2 | 13 | 0.000146 | 0.001247 |
| 539 | HEART VALVE FORMATION | 2 | 13 | 0.000146 | 0.001247 |
| 540 | RESPONSE TO HEPATOCYTE GROWTH FACTOR | 2 | 13 | 0.000146 | 0.001247 |
| 541 | POSITIVE REGULATION OF CYTOKINE PRODUCTION | 5 | 370 | 0.000146 | 0.001247 |
| 542 | NEUROBLAST DIVISION | 2 | 13 | 0.000146 | 0.001247 |
| 543 | GLIAL CELL FATE COMMITMENT | 2 | 13 | 0.000146 | 0.001247 |
| 544 | REGULATION OF BICELLULAR TIGHT JUNCTION ASSEMBLY | 2 | 13 | 0.000146 | 0.001247 |
| 545 | NEGATIVE REGULATION OF DNA DAMAGE RESPONSE SIGNAL TRANSDUCTION BY P53 CLASS MEDIATOR | 2 | 13 | 0.000146 | 0.001247 |
| 546 | REGULATION OF STEROID METABOLIC PROCESS | 3 | 74 | 0.000149 | 0.00127 |
| 547 | REGULATION OF STRESS ACTIVATED PROTEIN KINASE SIGNALING CASCADE | 4 | 197 | 0.0001553 | 0.001319 |
| 548 | BIOMINERAL TISSUE DEVELOPMENT | 3 | 75 | 0.0001551 | 0.001319 |
| 549 | PROTEIN COMPLEX SUBUNIT ORGANIZATION | 9 | 1527 | 0.0001567 | 0.001328 |
| 550 | REGULATION OF INTRACELLULAR PROTEIN TRANSPORT | 5 | 381 | 0.0001674 | 0.001416 |
| 551 | CELL MIGRATION INVOLVED IN HEART DEVELOPMENT | 2 | 14 | 0.0001702 | 0.001427 |
| 552 | SPECIFICATION OF ORGAN IDENTITY | 2 | 14 | 0.0001702 | 0.001427 |
| 553 | REGULATION OF GLOMERULUS DEVELOPMENT | 2 | 14 | 0.0001702 | 0.001427 |
| 554 | REGULATION OF PROTEIN KINASE C SIGNALING | 2 | 14 | 0.0001702 | 0.001427 |
| 555 | MIDGUT DEVELOPMENT | 2 | 14 | 0.0001702 | 0.001427 |
| 556 | NEGATIVE REGULATION OF PROTEIN MODIFICATION PROCESS | 6 | 616 | 0.0001759 | 0.001472 |
| 557 | REGULATION OF LEUKOCYTE APOPTOTIC PROCESS | 3 | 79 | 0.000181 | 0.001506 |
| 558 | REGULATION OF SYMBIOSIS ENCOMPASSING MUTUALISM THROUGH PARASITISM | 4 | 205 | 0.000181 | 0.001506 |
| 559 | NEURON PROJECTION GUIDANCE | 4 | 205 | 0.000181 | 0.001506 |
| 560 | NEGATIVE REGULATION OF LIPID METABOLIC PROCESS | 3 | 80 | 0.0001878 | 0.001561 |
| 561 | CELL PROJECTION ORGANIZATION | 7 | 902 | 0.0001909 | 0.001584 |
| 562 | POSITIVE REGULATION OF ENDOTHELIAL CELL DIFFERENTIATION | 2 | 15 | 0.0001962 | 0.001613 |
| 563 | METANEPHROS DEVELOPMENT | 3 | 81 | 0.0001949 | 0.001613 |
| 564 | OTIC VESICLE DEVELOPMENT | 2 | 15 | 0.0001962 | 0.001613 |
| 565 | POSITIVE REGULATION OF T CELL APOPTOTIC PROCESS | 2 | 15 | 0.0001962 | 0.001613 |
| 566 | EPITHELIAL CELL FATE COMMITMENT | 2 | 15 | 0.0001962 | 0.001613 |
| 567 | RESPONSE TO MECHANICAL STIMULUS | 4 | 210 | 0.0001985 | 0.001629 |
| 568 | NEGATIVE REGULATION OF CYTOKINE PRODUCTION | 4 | 211 | 0.0002021 | 0.001656 |
| 569 | CELLULAR MACROMOLECULE LOCALIZATION | 8 | 1234 | 0.0002099 | 0.001717 |
| 570 | TISSUE MIGRATION | 3 | 84 | 0.000217 | 0.001772 |
| 571 | RESPONSE TO PEPTIDE | 5 | 404 | 0.0002198 | 0.001788 |
| 572 | SIGNAL TRANSDUCTION BY PROTEIN PHOSPHORYLATION | 5 | 404 | 0.0002198 | 0.001788 |
| 573 | REGULATION OF STRIATED MUSCLE CELL DIFFERENTIATION | 3 | 85 | 0.0002248 | 0.001809 |
| 574 | ORGAN INDUCTION | 2 | 16 | 0.0002241 | 0.001809 |
| 575 | POSITIVE REGULATION OF CHROMATIN MODIFICATION | 3 | 85 | 0.0002248 | 0.001809 |
| 576 | APOPTOTIC PROCESS INVOLVED IN MORPHOGENESIS | 2 | 16 | 0.0002241 | 0.001809 |
| 577 | ATRIOVENTRICULAR VALVE MORPHOGENESIS | 2 | 16 | 0.0002241 | 0.001809 |
| 578 | BRANCHING INVOLVED IN SALIVARY GLAND MORPHOGENESIS | 2 | 16 | 0.0002241 | 0.001809 |
| 579 | NEGATIVE REGULATION OF KIDNEY DEVELOPMENT | 2 | 17 | 0.0002537 | 0.002015 |
| 580 | CELLULAR RESPONSE TO LITHIUM ION | 2 | 17 | 0.0002537 | 0.002015 |
| 581 | REGULATION OF RECEPTOR BINDING | 2 | 17 | 0.0002537 | 0.002015 |
| 582 | POSITIVE REGULATION OF EXTRACELLULAR MATRIX ORGANIZATION | 2 | 17 | 0.0002537 | 0.002015 |
| 583 | ESTABLISHMENT OF TISSUE POLARITY | 2 | 17 | 0.0002537 | 0.002015 |
| 584 | NEGATIVE REGULATION OF ANOIKIS | 2 | 17 | 0.0002537 | 0.002015 |
| 585 | REGULATION OF STEM CELL POPULATION MAINTENANCE | 2 | 17 | 0.0002537 | 0.002015 |
| 586 | NEGATIVE REGULATION OF CARDIAC MUSCLE TISSUE DEVELOPMENT | 2 | 17 | 0.0002537 | 0.002015 |
| 587 | CHROMATIN ORGANIZATION | 6 | 663 | 0.0002619 | 0.002076 |
| 588 | NEGATIVE REGULATION OF INTRINSIC APOPTOTIC SIGNALING PATHWAY | 3 | 90 | 0.0002661 | 0.002095 |
| 589 | MIDBRAIN DEVELOPMENT | 3 | 90 | 0.0002661 | 0.002095 |
| 590 | ENDOTHELIUM DEVELOPMENT | 3 | 90 | 0.0002661 | 0.002095 |
| 591 | REGULATION OF CELL MATRIX ADHESION | 3 | 90 | 0.0002661 | 0.002095 |
| 592 | INNER EAR MORPHOGENESIS | 3 | 92 | 0.0002839 | 0.002231 |
| 593 | REGULATION OF HORMONE BIOSYNTHETIC PROCESS | 2 | 18 | 0.0002852 | 0.002238 |
| 594 | POSITIVE REGULATION OF NEURON PROJECTION DEVELOPMENT | 4 | 232 | 0.0002902 | 0.002274 |
| 595 | REGULATION OF DNA BINDING | 3 | 93 | 0.0002931 | 0.002292 |
| 596 | REGULATION OF DNA BIOSYNTHETIC PROCESS | 3 | 94 | 0.0003025 | 0.002357 |
| 597 | NEURAL TUBE FORMATION | 3 | 94 | 0.0003025 | 0.002357 |
| 598 | NEGATIVE REGULATION OF CELLULAR COMPONENT ORGANIZATION | 6 | 684 | 0.0003097 | 0.00241 |
| 599 | NEGATIVE REGULATION OF INTRACELLULAR PROTEIN TRANSPORT | 3 | 95 | 0.000312 | 0.002416 |
| 600 | REGULATION OF TRANSCRIPTION FACTOR IMPORT INTO NUCLEUS | 3 | 95 | 0.000312 | 0.002416 |
| 601 | TRANSFORMING GROWTH FACTOR BETA RECEPTOR SIGNALING PATHWAY | 3 | 95 | 0.000312 | 0.002416 |
| 602 | REGULATION OF CARDIAC MUSCLE CELL DIFFERENTIATION | 2 | 19 | 0.0003185 | 0.002453 |
| 603 | ATRIOVENTRICULAR VALVE DEVELOPMENT | 2 | 19 | 0.0003185 | 0.002453 |
| 604 | POSITIVE REGULATION OF CHONDROCYTE DIFFERENTIATION | 2 | 19 | 0.0003185 | 0.002453 |
| 605 | MYELOID LEUKOCYTE DIFFERENTIATION | 3 | 96 | 0.0003218 | 0.002475 |
| 606 | NEGATIVE REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY | 3 | 98 | 0.0003419 | 0.002625 |
| 607 | POSITIVE REGULATION OF LYMPHOCYTE APOPTOTIC PROCESS | 2 | 20 | 0.0003536 | 0.002697 |
| 608 | NEGATIVE REGULATION OF BIOMINERAL TISSUE DEVELOPMENT | 2 | 20 | 0.0003536 | 0.002697 |
| 609 | TONGUE DEVELOPMENT | 2 | 20 | 0.0003536 | 0.002697 |
| 610 | POSITIVE REGULATION OF CELL SUBSTRATE ADHESION | 3 | 99 | 0.0003523 | 0.002697 |
| 611 | CELL DEATH | 7 | 1001 | 0.0003613 | 0.002751 |
| 612 | LIMBIC SYSTEM DEVELOPMENT | 3 | 100 | 0.0003628 | 0.002759 |
| 613 | POSITIVE REGULATION OF CELL CYCLE PROCESS | 4 | 247 | 0.0003682 | 0.002795 |
| 614 | KERATINOCYTE DIFFERENTIATION | 3 | 101 | 0.0003736 | 0.002831 |
| 615 | ECTODERM DEVELOPMENT | 2 | 21 | 0.0003904 | 0.002925 |
| 616 | CELL AGGREGATION | 2 | 21 | 0.0003904 | 0.002925 |
| 617 | APOPTOTIC PROCESS INVOLVED IN DEVELOPMENT | 2 | 21 | 0.0003904 | 0.002925 |
| 618 | POSITIVE REGULATION OF ADHERENS JUNCTION ORGANIZATION | 2 | 21 | 0.0003904 | 0.002925 |
| 619 | COCHLEA MORPHOGENESIS | 2 | 21 | 0.0003904 | 0.002925 |
| 620 | RESPONSE TO CYTOKINE | 6 | 714 | 0.0003897 | 0.002925 |
| 621 | CARTILAGE CONDENSATION | 2 | 21 | 0.0003904 | 0.002925 |
| 622 | REGULATION OF NEURON DEATH | 4 | 252 | 0.0003972 | 0.002971 |
| 623 | CEREBRAL CORTEX DEVELOPMENT | 3 | 105 | 0.0004187 | 0.003127 |
| 624 | REGULATION OF FAT CELL DIFFERENTIATION | 3 | 106 | 0.0004305 | 0.003184 |
| 625 | BETA CATENIN DESTRUCTION COMPLEX DISASSEMBLY | 2 | 22 | 0.0004291 | 0.003184 |
| 626 | MODULATION OF TRANSCRIPTION IN OTHER ORGANISM INVOLVED IN SYMBIOTIC INTERACTION | 2 | 22 | 0.0004291 | 0.003184 |
| 627 | FAT CELL DIFFERENTIATION | 3 | 106 | 0.0004305 | 0.003184 |
| 628 | PROTEIN LOCALIZATION TO CELL SURFACE | 2 | 22 | 0.0004291 | 0.003184 |
| 629 | POSITIVE REGULATION OF MESONEPHROS DEVELOPMENT | 2 | 22 | 0.0004291 | 0.003184 |
| 630 | REGULATION OF CATABOLIC PROCESS | 6 | 731 | 0.0004418 | 0.003263 |
| 631 | RESPONSE TO RETINOIC ACID | 3 | 107 | 0.0004425 | 0.003263 |
| 632 | REGULATION OF OSTEOBLAST PROLIFERATION | 2 | 23 | 0.0004696 | 0.003457 |
| 633 | CELLULAR RESPONSE TO EXTERNAL STIMULUS | 4 | 264 | 0.0004734 | 0.003475 |
| 634 | AGING | 4 | 264 | 0.0004734 | 0.003475 |
| 635 | REGULATION OF CYTOPLASMIC TRANSPORT | 5 | 481 | 0.0004901 | 0.003591 |
| 636 | POSITIVE REGULATION OF CELL JUNCTION ASSEMBLY | 2 | 24 | 0.0005118 | 0.003733 |
| 637 | NEGATIVE REGULATION OF MYOBLAST DIFFERENTIATION | 2 | 24 | 0.0005118 | 0.003733 |
| 638 | REGULATION OF ANOIKIS | 2 | 24 | 0.0005118 | 0.003733 |
| 639 | REGULATION OF CELL DIVISION | 4 | 272 | 0.0005298 | 0.003857 |
| 640 | NEGATIVE REGULATION OF RESPONSE TO EXTERNAL STIMULUS | 4 | 274 | 0.0005445 | 0.003959 |
| 641 | CELLULAR RESPONSE TO EPIDERMAL GROWTH FACTOR STIMULUS | 2 | 25 | 0.0005558 | 0.004035 |
| 642 | NEGATIVE REGULATION OF CYTOPLASMIC TRANSPORT | 3 | 117 | 0.0005745 | 0.004152 |
| 643 | SPECIFICATION OF SYMMETRY | 3 | 117 | 0.0005745 | 0.004152 |
| 644 | CELL CYCLE PROCESS | 7 | 1081 | 0.0005746 | 0.004152 |
| 645 | REGULATION OF PEPTIDYL SERINE PHOSPHORYLATION | 3 | 118 | 0.0005889 | 0.004248 |
| 646 | NEGATIVE REGULATION OF PROTEIN METABOLIC PROCESS | 7 | 1087 | 0.000594 | 0.004278 |
| 647 | DEVELOPMENTAL PROGRAMMED CELL DEATH | 2 | 26 | 0.0006016 | 0.004287 |
| 648 | NEGATIVE REGULATION OF STRIATED MUSCLE CELL DIFFERENTIATION | 2 | 26 | 0.0006016 | 0.004287 |
| 649 | REGULATION OF MESONEPHROS DEVELOPMENT | 2 | 26 | 0.0006016 | 0.004287 |
| 650 | MESODERMAL CELL DIFFERENTIATION | 2 | 26 | 0.0006016 | 0.004287 |
| 651 | REGULATION OF HORMONE METABOLIC PROCESS | 2 | 26 | 0.0006016 | 0.004287 |
| 652 | REGULATION OF P38MAPK CASCADE | 2 | 26 | 0.0006016 | 0.004287 |
| 653 | REGULATION OF CELL FATE COMMITMENT | 2 | 26 | 0.0006016 | 0.004287 |
| 654 | POSITIVE REGULATION OF NUCLEOCYTOPLASMIC TRANSPORT | 3 | 121 | 0.0006336 | 0.004508 |
| 655 | REGULATION OF ENDOTHELIAL CELL DIFFERENTIATION | 2 | 27 | 0.0006492 | 0.004584 |
| 656 | NEGATIVE REGULATION OF INTRINSIC APOPTOTIC SIGNALING PATHWAY IN RESPONSE TO DNA DAMAGE | 2 | 27 | 0.0006492 | 0.004584 |
| 657 | INFLAMMATORY RESPONSE TO ANTIGENIC STIMULUS | 2 | 27 | 0.0006492 | 0.004584 |
| 658 | NEGATIVE REGULATION OF SIGNAL TRANSDUCTION BY P53 CLASS MEDIATOR | 2 | 27 | 0.0006492 | 0.004584 |
| 659 | SUBSTRATE DEPENDENT CELL MIGRATION | 2 | 27 | 0.0006492 | 0.004584 |
| 660 | POSITIVE REGULATION OF LEUKOCYTE APOPTOTIC PROCESS | 2 | 28 | 0.0006985 | 0.004866 |
| 661 | GASTRULATION WITH MOUTH FORMING SECOND | 2 | 28 | 0.0006985 | 0.004866 |
| 662 | REGULATION OF DNA DAMAGE RESPONSE SIGNAL TRANSDUCTION BY P53 CLASS MEDIATOR | 2 | 28 | 0.0006985 | 0.004866 |
| 663 | AUDITORY RECEPTOR CELL DIFFERENTIATION | 2 | 28 | 0.0006985 | 0.004866 |
| 664 | STEROID HORMONE MEDIATED SIGNALING PATHWAY | 3 | 125 | 0.0006965 | 0.004866 |
| 665 | MORPHOGENESIS OF A POLARIZED EPITHELIUM | 2 | 28 | 0.0006985 | 0.004866 |
| 666 | RESPONSE TO GONADOTROPIN | 2 | 28 | 0.0006985 | 0.004866 |
| 667 | MAMMARY GLAND DUCT MORPHOGENESIS | 2 | 28 | 0.0006985 | 0.004866 |
| 668 | METANEPHROS MORPHOGENESIS | 2 | 28 | 0.0006985 | 0.004866 |
| 669 | CELLULAR RESPONSE TO OXYGEN CONTAINING COMPOUND | 6 | 799 | 0.0007069 | 0.004917 |
| 670 | REGULATION OF LIPID BIOSYNTHETIC PROCESS | 3 | 128 | 0.0007462 | 0.005182 |
| 671 | EMBRYONIC HINDLIMB MORPHOGENESIS | 2 | 29 | 0.0007496 | 0.005183 |
| 672 | REGULATION OF OLIGODENDROCYTE DIFFERENTIATION | 2 | 29 | 0.0007496 | 0.005183 |
| 673 | NEUROBLAST PROLIFERATION | 2 | 29 | 0.0007496 | 0.005183 |
| 674 | POSITIVE REGULATION OF PROTEIN LOCALIZATION TO NUCLEUS | 3 | 129 | 0.0007632 | 0.005261 |
| 675 | CELL JUNCTION ASSEMBLY | 3 | 129 | 0.0007632 | 0.005261 |
| 676 | PROTEIN LOCALIZATION TO CYTOSKELETON | 2 | 30 | 0.0008025 | 0.005499 |
| 677 | ESTABLISHMENT OR MAINTENANCE OF EPITHELIAL CELL APICAL BASAL POLARITY | 2 | 30 | 0.0008025 | 0.005499 |
| 678 | RESPONSE TO EPIDERMAL GROWTH FACTOR | 2 | 30 | 0.0008025 | 0.005499 |
| 679 | NEGATIVE REGULATION OF MUSCLE CELL APOPTOTIC PROCESS | 2 | 30 | 0.0008025 | 0.005499 |
| 680 | POSITIVE REGULATION OF CELL DIVISION | 3 | 132 | 0.0008159 | 0.005583 |
| 681 | POSITIVE REGULATION OF VASCULATURE DEVELOPMENT | 3 | 133 | 0.0008339 | 0.005685 |
| 682 | NEGATIVE REGULATION OF PHOSPHORUS METABOLIC PROCESS | 5 | 541 | 0.0008344 | 0.005685 |
| 683 | NEGATIVE REGULATION OF PHOSPHATE METABOLIC PROCESS | 5 | 541 | 0.0008344 | 0.005685 |
| 684 | CARDIAC ATRIUM DEVELOPMENT | 2 | 31 | 0.0008571 | 0.005825 |
| 685 | NEGATIVE REGULATION OF CATALYTIC ACTIVITY | 6 | 829 | 0.0008575 | 0.005825 |
| 686 | NEGATIVE REGULATION OF CELLULAR PROTEIN LOCALIZATION | 3 | 135 | 0.0008708 | 0.005898 |
| 687 | POSITIVE REGULATION OF STRESS ACTIVATED PROTEIN KINASE SIGNALING CASCADE | 3 | 135 | 0.0008708 | 0.005898 |
| 688 | HINDBRAIN DEVELOPMENT | 3 | 137 | 0.0009087 | 0.006107 |
| 689 | POSITIVE REGULATION OF GLIAL CELL DIFFERENTIATION | 2 | 32 | 0.0009134 | 0.006107 |
| 690 | POSITIVE REGULATION OF BMP SIGNALING PATHWAY | 2 | 32 | 0.0009134 | 0.006107 |
| 691 | REGULATION OF HEMOPOIESIS | 4 | 314 | 0.0009063 | 0.006107 |
| 692 | PATTERNING OF BLOOD VESSELS | 2 | 32 | 0.0009134 | 0.006107 |
| 693 | CELLULAR RESPONSE TO HORMONE STIMULUS | 5 | 552 | 0.0009134 | 0.006107 |
| 694 | POSITIVE REGULATION OF EPIDERMIS DEVELOPMENT | 2 | 32 | 0.0009134 | 0.006107 |
| 695 | BLOOD VESSEL REMODELING | 2 | 32 | 0.0009134 | 0.006107 |
| 696 | ACTIVATION OF MAPK ACTIVITY | 3 | 137 | 0.0009087 | 0.006107 |
| 697 | PEPTIDYL AMINO ACID MODIFICATION | 6 | 841 | 0.0009242 | 0.00617 |
| 698 | REGULATION OF CELL PROJECTION ORGANIZATION | 5 | 558 | 0.0009587 | 0.006391 |
| 699 | REGULATION OF MYELINATION | 2 | 33 | 0.0009715 | 0.006458 |
| 700 | POSITIVE REGULATION OF RESPONSE TO CYTOKINE STIMULUS | 2 | 33 | 0.0009715 | 0.006458 |
| 701 | ESTABLISHMENT OR MAINTENANCE OF CELL POLARITY | 3 | 141 | 0.0009875 | 0.006555 |
| 702 | FOREBRAIN NEURON DEVELOPMENT | 2 | 34 | 0.001031 | 0.0068 |
| 703 | RESPONSE TO NITROGEN COMPOUND | 6 | 859 | 0.001032 | 0.0068 |
| 704 | LUNG EPITHELIUM DEVELOPMENT | 2 | 34 | 0.001031 | 0.0068 |
| 705 | REGULATION OF INTRINSIC APOPTOTIC SIGNALING PATHWAY IN RESPONSE TO DNA DAMAGE | 2 | 34 | 0.001031 | 0.0068 |
| 706 | NEGATIVE REGULATION OF INTRACELLULAR TRANSPORT | 3 | 143 | 0.001029 | 0.0068 |
| 707 | CELLULAR RESPONSE TO STRESS | 8 | 1565 | 0.001037 | 0.006827 |
| 708 | REGULATION OF INTRINSIC APOPTOTIC SIGNALING PATHWAY | 3 | 145 | 0.001071 | 0.007036 |
| 709 | NEGATIVE REGULATION OF RESPONSE TO OXIDATIVE STRESS | 2 | 35 | 0.001093 | 0.007152 |
| 710 | NEGATIVE REGULATION OF CELLULAR RESPONSE TO OXIDATIVE STRESS | 2 | 35 | 0.001093 | 0.007152 |
| 711 | REGULATION OF RESPONSE TO REACTIVE OXYGEN SPECIES | 2 | 35 | 0.001093 | 0.007152 |
| 712 | PEPTIDYL SERINE MODIFICATION | 3 | 148 | 0.001136 | 0.007422 |
| 713 | REGULATION OF TRANSCRIPTION REGULATORY REGION DNA BINDING | 2 | 36 | 0.001156 | 0.007535 |
| 714 | POSITIVE CHEMOTAXIS | 2 | 36 | 0.001156 | 0.007535 |
| 715 | POSITIVE REGULATION OF CHROMOSOME ORGANIZATION | 3 | 150 | 0.001181 | 0.007672 |
| 716 | CHROMATIN REMODELING | 3 | 150 | 0.001181 | 0.007672 |
| 717 | SECRETION | 5 | 588 | 0.001211 | 0.007849 |
| 718 | RESPONSE TO BIOTIC STIMULUS | 6 | 886 | 0.001211 | 0.007849 |
| 719 | HINDLIMB MORPHOGENESIS | 2 | 37 | 0.001221 | 0.00787 |
| 720 | GLIAL CELL MIGRATION | 2 | 37 | 0.001221 | 0.00787 |
| 721 | ESTABLISHMENT OR MAINTENANCE OF APICAL BASAL CELL POLARITY | 2 | 37 | 0.001221 | 0.00787 |
| 722 | ESTABLISHMENT OR MAINTENANCE OF BIPOLAR CELL POLARITY | 2 | 37 | 0.001221 | 0.00787 |
| 723 | REGULATION OF CHROMATIN ORGANIZATION | 3 | 152 | 0.001226 | 0.007893 |
| 724 | MULTI ORGANISM REPRODUCTIVE PROCESS | 6 | 891 | 0.001247 | 0.008013 |
| 725 | TRANSCRIPTION INITIATION FROM RNA POLYMERASE II PROMOTER | 3 | 153 | 0.00125 | 0.008021 |
| 726 | POSITIVE REGULATION OF BIOMINERAL TISSUE DEVELOPMENT | 2 | 38 | 0.001288 | 0.008243 |
| 727 | CYTOKINE SECRETION | 2 | 38 | 0.001288 | 0.008243 |
| 728 | BONE DEVELOPMENT | 3 | 156 | 0.001322 | 0.008447 |
| 729 | POSITIVE REGULATION OF VIRAL TRANSCRIPTION | 2 | 39 | 0.001356 | 0.008599 |
| 730 | NEGATIVE REGULATION OF TRANSCRIPTION FACTOR IMPORT INTO NUCLEUS | 2 | 39 | 0.001356 | 0.008599 |
| 731 | POSITIVE REGULATION OF HYDROLASE ACTIVITY | 6 | 905 | 0.001351 | 0.008599 |
| 732 | REGULATION OF CELL ADHESION MEDIATED BY INTEGRIN | 2 | 39 | 0.001356 | 0.008599 |
| 733 | ASTROCYTE DIFFERENTIATION | 2 | 39 | 0.001356 | 0.008599 |
| 734 | COCHLEA DEVELOPMENT | 2 | 39 | 0.001356 | 0.008599 |
| 735 | HORMONE MEDIATED SIGNALING PATHWAY | 3 | 158 | 0.001371 | 0.008678 |
| 736 | CELLULAR RESPONSE TO CYTOKINE STIMULUS | 5 | 606 | 0.001385 | 0.008754 |
| 737 | MAMMARY GLAND MORPHOGENESIS | 2 | 40 | 0.001427 | 0.00897 |
| 738 | ENDOCRINE PANCREAS DEVELOPMENT | 2 | 40 | 0.001427 | 0.00897 |
| 739 | ENDODERMAL CELL DIFFERENTIATION | 2 | 40 | 0.001427 | 0.00897 |
| 740 | NEGATIVE REGULATION OF OSTEOBLAST DIFFERENTIATION | 2 | 40 | 0.001427 | 0.00897 |
| 741 | CELL CHEMOTAXIS | 3 | 162 | 0.001473 | 0.009248 |
| 742 | CELLULAR RESPONSE TO BIOTIC STIMULUS | 3 | 163 | 0.001499 | 0.009374 |
| 743 | POSITIVE REGULATION OF KIDNEY DEVELOPMENT | 2 | 41 | 0.001498 | 0.009374 |
| 744 | LUNG ALVEOLUS DEVELOPMENT | 2 | 41 | 0.001498 | 0.009374 |
| 745 | POSITIVE REGULATION OF CELLULAR PROTEIN LOCALIZATION | 4 | 360 | 0.001502 | 0.009381 |
| 746 | REGULATION OF INTRACELLULAR TRANSPORT | 5 | 621 | 0.001543 | 0.009623 |
| 747 | CENTRAL NERVOUS SYSTEM NEURON DIFFERENTIATION | 3 | 166 | 0.001579 | 0.009838 |
| 748 | POSITIVE REGULATION OF TRANSPORT | 6 | 936 | 0.001606 | 0.009991 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | NUCLEIC ACID BINDING TRANSCRIPTION FACTOR ACTIVITY | 13 | 1199 | 1.947e-09 | 9.045e-07 |
| 2 | MACROMOLECULAR COMPLEX BINDING | 14 | 1399 | 9.958e-10 | 9.045e-07 |
| 3 | CHROMATIN BINDING | 9 | 435 | 4.802e-09 | 1.487e-06 |
| 4 | TRANSFORMING GROWTH FACTOR BETA RECEPTOR BINDING | 5 | 50 | 7.482e-09 | 1.738e-06 |
| 5 | RECEPTOR BINDING | 13 | 1476 | 2.395e-08 | 4.451e-06 |
| 6 | SMAD BINDING | 5 | 72 | 4.838e-08 | 7.491e-06 |
| 7 | SEQUENCE SPECIFIC DNA BINDING | 11 | 1037 | 6.566e-08 | 8.715e-06 |
| 8 | REGULATORY REGION NUCLEIC ACID BINDING | 10 | 818 | 8.324e-08 | 9.666e-06 |
| 9 | TRANSCRIPTION FACTOR ACTIVITY RNA POLYMERASE II DISTAL ENHANCER SEQUENCE SPECIFIC BINDING | 5 | 90 | 1.494e-07 | 1.542e-05 |
| 10 | TRANSCRIPTION FACTOR ACTIVITY RNA POLYMERASE II CORE PROMOTER PROXIMAL REGION SEQUENCE SPECIFIC BINDING | 7 | 328 | 2.636e-07 | 2.449e-05 |
| 11 | TRANSCRIPTION FACTOR BINDING | 8 | 524 | 4.122e-07 | 3.481e-05 |
| 12 | RNA POLYMERASE II TRANSCRIPTION FACTOR ACTIVITY SEQUENCE SPECIFIC DNA BINDING | 8 | 629 | 1.63e-06 | 0.0001262 |
| 13 | PROTEIN KINASE ACTIVITY | 8 | 640 | 1.855e-06 | 0.0001326 |
| 14 | TRANSCRIPTIONAL ACTIVATOR ACTIVITY RNA POLYMERASE II TRANSCRIPTION REGULATORY REGION SEQUENCE SPECIFIC BINDING | 6 | 315 | 4.094e-06 | 0.0002717 |
| 15 | KINASE ACTIVITY | 8 | 842 | 1.4e-05 | 0.0008136 |
| 16 | TRANSCRIPTIONAL ACTIVATOR ACTIVITY RNA POLYMERASE II CORE PROMOTER PROXIMAL REGION SEQUENCE SPECIFIC BINDING | 5 | 226 | 1.401e-05 | 0.0008136 |
| 17 | KINASE BINDING | 7 | 606 | 1.54e-05 | 0.0008415 |
| 18 | CYTOKINE RECEPTOR BINDING | 5 | 271 | 3.352e-05 | 0.00173 |
| 19 | PROTEIN HETERODIMERIZATION ACTIVITY | 6 | 468 | 3.867e-05 | 0.001891 |
| 20 | TRANSFERASE ACTIVITY TRANSFERRING PHOSPHORUS CONTAINING GROUPS | 8 | 992 | 4.552e-05 | 0.002114 |
| 21 | CORE PROMOTER BINDING | 4 | 152 | 5.696e-05 | 0.00252 |
| 22 | GROWTH FACTOR ACTIVITY | 4 | 160 | 6.954e-05 | 0.002805 |
| 23 | DOUBLE STRANDED DNA BINDING | 7 | 764 | 6.771e-05 | 0.002805 |
| 24 | ENZYME BINDING | 10 | 1737 | 7.245e-05 | 0.002805 |
| 25 | I SMAD BINDING | 2 | 11 | 0.0001031 | 0.003833 |
| 26 | PROTEIN DIMERIZATION ACTIVITY | 8 | 1149 | 0.0001281 | 0.004577 |
| 27 | TRANSCRIPTION FACTOR ACTIVITY PROTEIN BINDING | 6 | 588 | 0.0001365 | 0.004698 |
| 28 | TRANSMEMBRANE RECEPTOR PROTEIN KINASE ACTIVITY | 3 | 81 | 0.0001949 | 0.005524 |
| 29 | C2H2 ZINC FINGER DOMAIN BINDING | 2 | 14 | 0.0001702 | 0.005524 |
| 30 | RECEPTOR SERINE THREONINE KINASE BINDING | 2 | 15 | 0.0001962 | 0.005524 |
| 31 | PROTEIN KINASE A CATALYTIC SUBUNIT BINDING | 2 | 15 | 0.0001962 | 0.005524 |
| 32 | OXIDOREDUCTASE ACTIVITY ACTING ON THE CH NH2 GROUP OF DONORS OXYGEN AS ACCEPTOR | 2 | 15 | 0.0001962 | 0.005524 |
| 33 | PROTEIN DOMAIN SPECIFIC BINDING | 6 | 624 | 0.0001887 | 0.005524 |
| 34 | BETA CATENIN BINDING | 3 | 84 | 0.000217 | 0.00593 |
| 35 | TRANSFORMING GROWTH FACTOR BETA BINDING | 2 | 16 | 0.0002241 | 0.005948 |
| 36 | PROTEIN COMPLEX BINDING | 7 | 935 | 0.0002382 | 0.005981 |
| 37 | CYTOKINE ACTIVITY | 4 | 219 | 0.000233 | 0.005981 |
| 38 | TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE ACTIVITY | 2 | 17 | 0.0002537 | 0.006203 |
| 39 | CYTOKINE BINDING | 3 | 92 | 0.0002839 | 0.006763 |
| 40 | OXIDOREDUCTASE ACTIVITY ACTING ON THE CH NH2 GROUP OF DONORS | 2 | 19 | 0.0003185 | 0.007397 |
| 41 | PROTEIN SERINE THREONINE KINASE ACTIVITY | 5 | 445 | 0.0003433 | 0.007779 |
| 42 | DNA BINDING BENDING | 2 | 20 | 0.0003536 | 0.00782 |
| 43 | RNA POLYMERASE II TRANSCRIPTION FACTOR BINDING | 3 | 104 | 0.0004071 | 0.008795 |
| 44 | TRANSCRIPTIONAL REPRESSOR ACTIVITY RNA POLYMERASE II CORE PROMOTER PROXIMAL REGION SEQUENCE SPECIFIC BINDING | 3 | 105 | 0.0004187 | 0.00884 |
| 45 | IONOTROPIC GLUTAMATE RECEPTOR BINDING | 2 | 23 | 0.0004696 | 0.009483 |
| 46 | FIBROBLAST GROWTH FACTOR BINDING | 2 | 23 | 0.0004696 | 0.009483 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | TRANSCRIPTION FACTOR COMPLEX | 8 | 298 | 5.307e-09 | 3.099e-06 |
| 2 | NUCLEAR TRANSCRIPTION FACTOR COMPLEX | 6 | 127 | 1.956e-08 | 5.711e-06 |
| 3 | RNA POLYMERASE II TRANSCRIPTION FACTOR COMPLEX | 5 | 101 | 2.664e-07 | 5.185e-05 |
| 4 | EXTRACELLULAR MATRIX | 7 | 426 | 1.523e-06 | 0.0002223 |
| 5 | EXTRACELLULAR SPACE | 10 | 1376 | 9.586e-06 | 0.00112 |
Over-represented Pathway
| Num | Pathway | Pathview | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|---|
| 1 | hsa04390_Hippo_signaling_pathway | 10 | 154 | 6.353e-15 | 1.144e-12 | |
| 2 | hsa04340_Hedgehog_signaling_pathway | 5 | 56 | 1.341e-08 | 1.207e-06 | |
| 3 | hsa04520_Adherens_junction | 5 | 73 | 5.189e-08 | 3.113e-06 | |
| 4 | hsa04350_TGF.beta_signaling_pathway | 5 | 85 | 1.12e-07 | 5.041e-06 | |
| 5 | hsa04916_Melanogenesis | 5 | 101 | 2.664e-07 | 9.589e-06 | |
| 6 | hsa04310_Wnt_signaling_pathway | 5 | 151 | 1.961e-06 | 5.883e-05 | |
| 7 | hsa04330_Notch_signaling_pathway | 2 | 47 | 0.001965 | 0.05053 | |
| 8 | hsa04510_Focal_adhesion | 3 | 200 | 0.002684 | 0.05599 | |
| 9 | hsa04144_Endocytosis | 3 | 203 | 0.002799 | 0.05599 | |
| 10 | hsa04010_MAPK_signaling_pathway | 3 | 268 | 0.006085 | 0.1095 | |
| 11 | hsa04151_PI3K_AKT_signaling_pathway | 3 | 351 | 0.01269 | 0.1908 | |
| 12 | hsa04110_Cell_cycle | 2 | 128 | 0.01378 | 0.1908 | |
| 13 | hsa04380_Osteoclast_differentiation | 2 | 128 | 0.01378 | 0.1908 | |
| 14 | hsa04014_Ras_signaling_pathway | 2 | 236 | 0.04286 | 0.5143 |

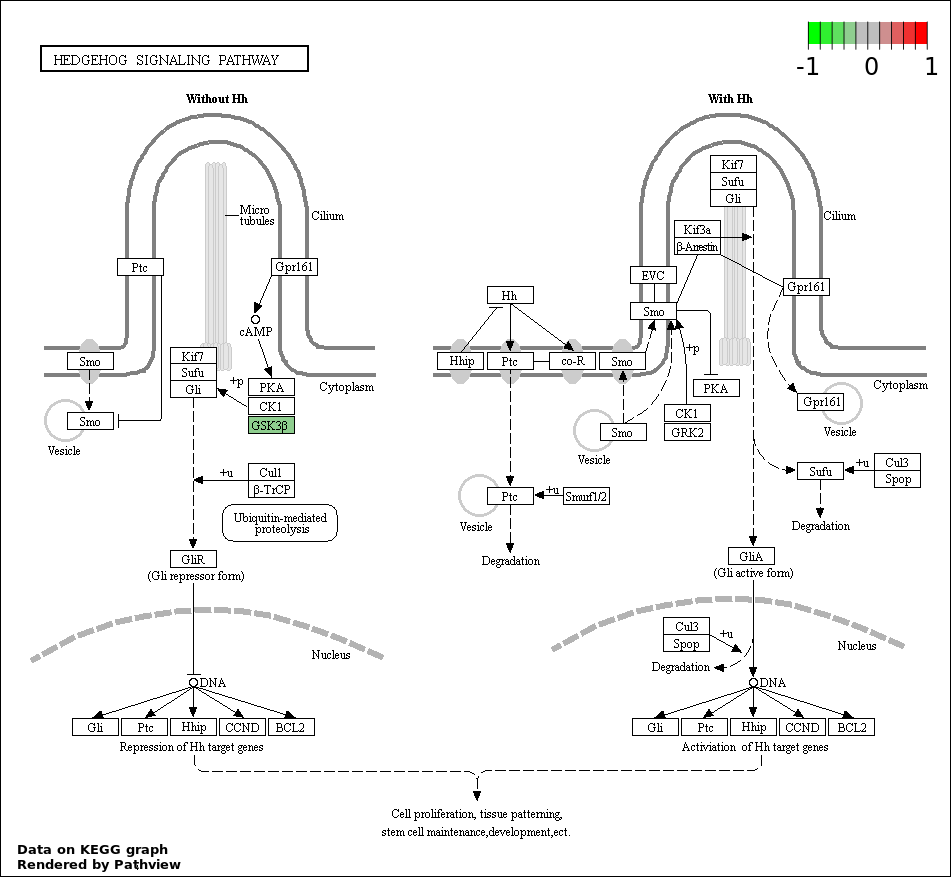


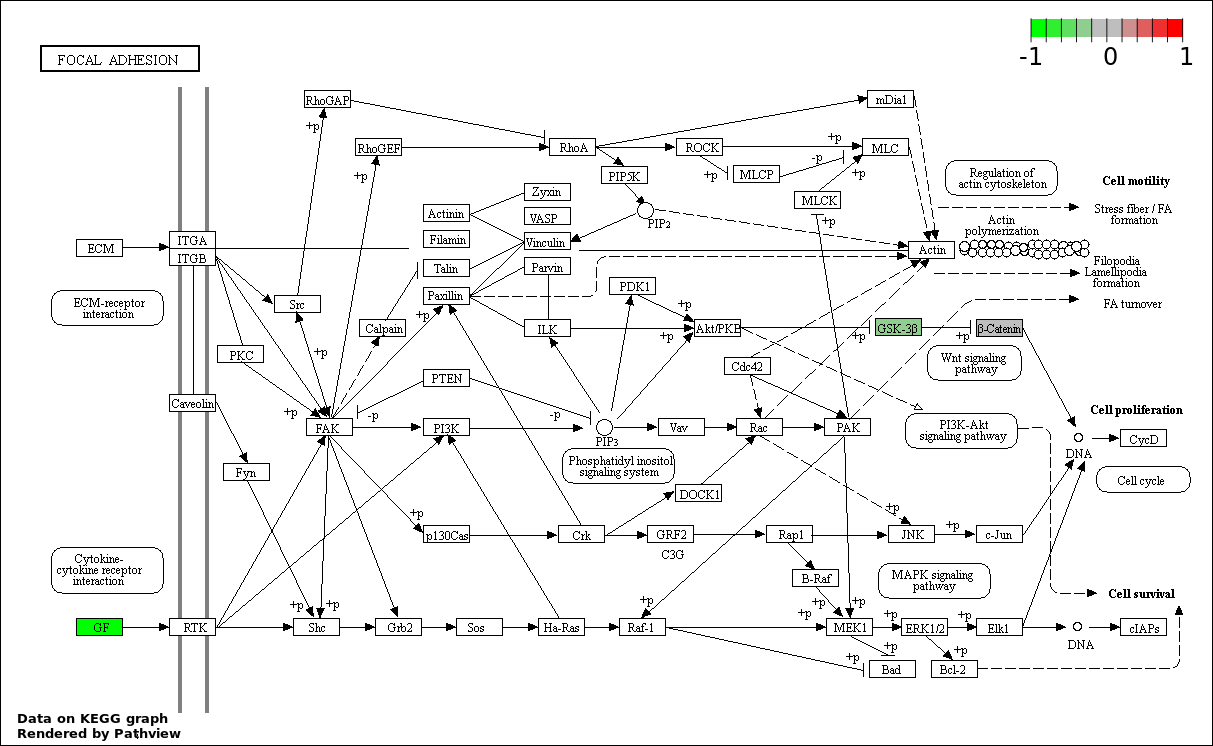





lncRNA-mediated sponge
| Num | lncRNA | miRNAs | miRNAs count | Gene | Sponge regulatory network | lncRNA log2FC | lncRNA pvalue | Gene log2FC | Gene pvalue | lncRNA-gene Pearson correlation |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | LINC00968 |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-339-5p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-625-5p | 16 | TGFBR3 | Sponge network | -4.19 | 0 | -3.002 | 0 | 0.578 |
| 2 | FENDRR |
hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-3607-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-590-3p;hsa-miR-7-1-3p;hsa-miR-98-5p | 13 | TGFBR3 | Sponge network | -4.222 | 0 | -3.002 | 0 | 0.57 |
| 3 | LINC00702 |
hsa-miR-130b-3p;hsa-miR-142-3p;hsa-miR-182-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200c-3p;hsa-miR-301a-3p;hsa-miR-454-3p;hsa-miR-590-3p;hsa-miR-96-5p | 10 | FOXF2 | Sponge network | -2.856 | 0 | -1.7 | 0 | 0.554 |
| 4 | RP11-389C8.2 |
hsa-miR-107;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-429;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-7-1-3p | 12 | TGFBR3 | Sponge network | -2.039 | 0 | -3.002 | 0 | 0.55 |
| 5 | AC109642.1 |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-424-5p;hsa-miR-590-3p;hsa-miR-98-5p | 13 | TGFBR3 | Sponge network | -2.791 | 0 | -3.002 | 0 | 0.55 |
| 6 | RP11-401P9.4 | hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-590-5p;hsa-miR-98-5p | 10 | TGFBR3 | Sponge network | -3.04 | 0 | -3.002 | 0 | 0.544 |
| 7 | MAGI2-AS3 |
hsa-miR-130b-3p;hsa-miR-182-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200c-3p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-454-3p;hsa-miR-590-3p;hsa-miR-9-5p | 10 | FOXF2 | Sponge network | -1.892 | 0 | -1.7 | 0 | 0.539 |
| 8 | TBX5-AS1 |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-450b-5p;hsa-miR-590-5p;hsa-miR-7-1-3p | 14 | TGFBR3 | Sponge network | -2.108 | 0 | -3.002 | 0 | 0.539 |
| 9 | LINC00702 |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-30e-5p;hsa-miR-335-3p;hsa-miR-339-5p;hsa-miR-34a-5p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-625-5p;hsa-miR-7-1-3p | 21 | TGFBR3 | Sponge network | -2.856 | 0 | -3.002 | 0 | 0.52 |
| 10 | MAGI2-AS3 |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-429;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-625-5p;hsa-miR-7-1-3p | 16 | TGFBR3 | Sponge network | -1.892 | 0 | -3.002 | 0 | 0.507 |
| 11 | RP11-1024P17.1 |
hsa-miR-107;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-429;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-625-5p | 14 | TGFBR3 | Sponge network | -2.062 | 0 | -3.002 | 0 | 0.502 |
| 12 | AF131215.2 | hsa-miR-107;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-589-3p;hsa-miR-7-1-3p;hsa-miR-98-5p | 10 | TGFBR3 | Sponge network | -2.09 | 0 | -3.002 | 0 | 0.483 |
| 13 | GAS6-AS2 |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-429;hsa-miR-590-5p;hsa-miR-7-1-3p | 13 | TGFBR3 | Sponge network | -1.761 | 0 | -3.002 | 0 | 0.457 |
| 14 | CTD-2013N24.2 |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-5p;hsa-miR-181b-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-30e-5p;hsa-miR-335-3p;hsa-miR-429;hsa-miR-590-3p;hsa-miR-625-5p;hsa-miR-7-1-3p | 15 | TGFBR3 | Sponge network | -1.745 | 0 | -3.002 | 0 | 0.448 |
| 15 | CASC2 |
hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-7-1-3p | 10 | TGFBR3 | Sponge network | -1.086 | 0 | -3.002 | 0 | 0.438 |
| 16 | AC004947.2 |
hsa-miR-16-2-3p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-335-3p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-7-1-3p | 11 | TGFBR3 | Sponge network | -3.94 | 0 | -3.002 | 0 | 0.433 |
| 17 | RP11-456K23.1 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-625-5p | 14 | TGFBR3 | Sponge network | -1.488 | 0 | -3.002 | 0 | 0.43 |
| 18 | RP11-354E11.2 |
hsa-miR-107;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-590-3p | 11 | TGFBR3 | Sponge network | -2.138 | 0 | -3.002 | 0 | 0.425 |
| 19 | LINC00702 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-454-3p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p | 18 | TGFB2 | Sponge network | -2.856 | 0 | -1.547 | 1.0E-5 | 0.413 |
| 20 | MIR497HG |
hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-339-5p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-625-5p;hsa-miR-7-1-3p | 17 | TGFBR3 | Sponge network | -2.142 | 0 | -3.002 | 0 | 0.404 |
| 21 | WDFY3-AS2 |
hsa-miR-107;hsa-miR-16-2-3p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-7-1-3p | 12 | TGFBR3 | Sponge network | -1.297 | 0 | -3.002 | 0 | 0.403 |
| 22 | DIO3OS | hsa-miR-107;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-21-5p;hsa-miR-424-5p;hsa-miR-589-3p;hsa-miR-625-5p;hsa-miR-98-5p | 10 | TGFBR3 | Sponge network | -1.936 | 0.00085 | -3.002 | 0 | 0.399 |
| 23 | RP11-378A13.1 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-590-3p | 10 | TGFBR3 | Sponge network | -1.713 | 0 | -3.002 | 0 | 0.392 |
| 24 | CTD-2013N24.2 |
hsa-miR-130b-3p;hsa-miR-142-3p;hsa-miR-182-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200c-3p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-454-3p;hsa-miR-590-3p | 10 | FOXF2 | Sponge network | -1.745 | 0 | -1.7 | 0 | 0.388 |
| 25 | AF131215.9 | hsa-miR-107;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-590-3p | 10 | TGFBR3 | Sponge network | -1.808 | 0 | -3.002 | 0 | 0.384 |
| 26 | AC079630.4 | hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-3607-3p;hsa-miR-424-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-625-5p;hsa-miR-7-1-3p | 10 | TGFBR3 | Sponge network | -3.758 | 0 | -3.002 | 0 | 0.377 |
| 27 | AC011899.9 |
hsa-miR-107;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-30e-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-590-3p;hsa-miR-7-1-3p | 13 | TGFBR3 | Sponge network | -2.611 | 0 | -3.002 | 0 | 0.372 |
| 28 | SH3RF3-AS1 | hsa-miR-107;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-18a-5p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-30e-5p;hsa-miR-339-5p;hsa-miR-590-5p;hsa-miR-7-1-3p | 13 | TGFBR3 | Sponge network | -1.583 | 0 | -3.002 | 0 | 0.37 |
| 29 | GAS6-AS2 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-590-5p | 10 | TGFB2 | Sponge network | -1.761 | 0 | -1.547 | 1.0E-5 | 0.369 |
| 30 | RP11-1024P17.1 |
hsa-miR-130b-3p;hsa-miR-142-3p;hsa-miR-182-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200c-3p;hsa-miR-301a-3p;hsa-miR-429;hsa-miR-590-3p;hsa-miR-9-5p | 10 | FOXF2 | Sponge network | -2.062 | 0 | -1.7 | 0 | 0.358 |
| 31 | RP11-354E11.2 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-590-3p | 11 | TGFB2 | Sponge network | -2.138 | 0 | -1.547 | 1.0E-5 | 0.356 |
| 32 | RP11-1024P17.1 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-335-3p;hsa-miR-589-3p;hsa-miR-590-3p | 14 | TGFB2 | Sponge network | -2.062 | 0 | -1.547 | 1.0E-5 | 0.354 |
| 33 | LINC00472 | hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-335-3p | 10 | TGFB2 | Sponge network | -2.952 | 0 | -1.547 | 1.0E-5 | 0.351 |
| 34 | LINC00261 |
hsa-miR-16-2-3p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-7-1-3p | 12 | TGFBR3 | Sponge network | -2.566 | 0.00025 | -3.002 | 0 | 0.346 |
| 35 | AC109642.1 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-590-3p | 12 | TGFB2 | Sponge network | -2.791 | 0 | -1.547 | 1.0E-5 | 0.343 |
| 36 | MAGI2-AS3 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-454-3p;hsa-miR-589-3p;hsa-miR-590-3p | 16 | TGFB2 | Sponge network | -1.892 | 0 | -1.547 | 1.0E-5 | 0.342 |
| 37 | AC007879.7 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-miR-129-5p;hsa-miR-195-5p;hsa-miR-204-5p;hsa-miR-29b-2-5p;hsa-miR-30d-3p;hsa-miR-326 | 10 | HMGA2 | Sponge network | 2.583 | 0 | 3.122 | 0.00011 | 0.34 |
| 38 | NR2F1-AS1 |
hsa-miR-141-3p;hsa-miR-15b-3p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-186-5p;hsa-miR-330-5p;hsa-miR-421;hsa-miR-582-5p;hsa-miR-590-3p;hsa-miR-7-1-3p;hsa-miR-92a-3p | 11 | WNT5A | Sponge network | -0.427 | 0.1559 | -0.372 | 0.19306 | 0.336 |
| 39 | CTD-2013N24.2 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-454-3p;hsa-miR-590-3p | 14 | TGFB2 | Sponge network | -1.745 | 0 | -1.547 | 1.0E-5 | 0.334 |
| 40 | RP13-463N16.6 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-let-7g-5p;hsa-miR-204-5p;hsa-miR-29b-2-5p;hsa-miR-326;hsa-miR-362-3p;hsa-miR-664a-3p | 10 | HMGA2 | Sponge network | 6.976 | 0 | 3.122 | 0.00011 | 0.332 |
| 41 | CTD-2587H24.5 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7g-5p;hsa-miR-195-3p;hsa-miR-195-5p;hsa-miR-204-5p;hsa-miR-29b-2-5p;hsa-miR-30d-3p;hsa-miR-326;hsa-miR-486-5p;hsa-miR-664a-5p | 12 | HMGA2 | Sponge network | 3.545 | 0.00306 | 3.122 | 0.00011 | 0.326 |
| 42 | TBX5-AS1 |
hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-15b-3p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-186-5p;hsa-miR-29a-5p;hsa-miR-330-5p;hsa-miR-421;hsa-miR-576-5p;hsa-miR-7-1-3p;hsa-miR-92a-3p | 12 | WNT5A | Sponge network | -2.108 | 0 | -0.372 | 0.19306 | 0.323 |
| 43 | MAGI2-AS3 |
hsa-miR-141-3p;hsa-miR-15b-3p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-186-5p;hsa-miR-29a-5p;hsa-miR-330-5p;hsa-miR-421;hsa-miR-576-5p;hsa-miR-590-3p;hsa-miR-7-1-3p;hsa-miR-92a-3p | 12 | WNT5A | Sponge network | -1.892 | 0 | -0.372 | 0.19306 | 0.323 |
| 44 | TBX5-AS1 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-590-5p | 14 | TGFB2 | Sponge network | -2.108 | 0 | -1.547 | 1.0E-5 | 0.32 |
| 45 | AC007743.1 | hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-424-5p;hsa-miR-429;hsa-miR-590-5p;hsa-miR-7-1-3p | 10 | TGFBR3 | Sponge network | -2.595 | 0 | -3.002 | 0 | 0.314 |
| 46 | LINC00968 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-200a-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-590-3p;hsa-miR-590-5p | 13 | TGFB2 | Sponge network | -4.19 | 0 | -1.547 | 1.0E-5 | 0.308 |
| 47 | HHIP-AS1 | hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148b-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-330-5p;hsa-miR-335-3p;hsa-miR-590-3p | 10 | TGFB2 | Sponge network | -2.807 | 0 | -1.547 | 1.0E-5 | 0.297 |
| 48 | RP11-221J22.2 | hsa-let-7b-5p;hsa-let-7c-5p;hsa-miR-129-5p;hsa-miR-195-3p;hsa-miR-195-5p;hsa-miR-204-5p;hsa-miR-26b-5p;hsa-miR-29b-2-5p;hsa-miR-30d-3p;hsa-miR-664a-3p | 10 | HMGA2 | Sponge network | 0.704 | 0.25926 | 3.122 | 0.00011 | 0.29 |
| 49 | RP11-221J22.1 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-miR-129-5p;hsa-miR-195-3p;hsa-miR-195-5p;hsa-miR-204-5p;hsa-miR-26b-5p;hsa-miR-29b-2-5p;hsa-miR-30d-3p;hsa-miR-362-3p;hsa-miR-664a-3p | 12 | HMGA2 | Sponge network | 0.62 | 0.36599 | 3.122 | 0.00011 | 0.29 |
| 50 | RP11-57A1.1 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-miR-129-5p;hsa-miR-195-5p;hsa-miR-204-5p;hsa-miR-29b-2-5p;hsa-miR-30b-5p;hsa-miR-30c-5p;hsa-miR-30d-3p;hsa-miR-30d-5p;hsa-miR-664a-5p | 13 | HMGA2 | Sponge network | 3.979 | 0 | 3.122 | 0.00011 | 0.286 |
| 51 | MIR4435-1HG | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-miR-195-3p;hsa-miR-204-5p;hsa-miR-29b-2-5p;hsa-miR-30d-3p;hsa-miR-326;hsa-miR-664a-3p;hsa-miR-664a-5p | 10 | HMGA2 | Sponge network | 0.795 | 0.0006 | 3.122 | 0.00011 | 0.278 |
| 52 | LINC00511 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-miR-129-5p;hsa-miR-195-3p;hsa-miR-195-5p;hsa-miR-204-5p;hsa-miR-29b-2-5p;hsa-miR-30b-5p;hsa-miR-30c-5p;hsa-miR-30d-3p;hsa-miR-30d-5p;hsa-miR-326;hsa-miR-362-3p;hsa-miR-486-5p;hsa-miR-491-5p;hsa-miR-664a-5p | 17 | HMGA2 | Sponge network | 2.848 | 0 | 3.122 | 0.00011 | 0.276 |
| 53 | FENDRR |
hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-142-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-324-5p;hsa-miR-335-3p;hsa-miR-590-3p | 12 | TGFB2 | Sponge network | -4.222 | 0 | -1.547 | 1.0E-5 | 0.274 |
| 54 | RP11-389C8.2 |
hsa-miR-107;hsa-miR-130b-3p;hsa-miR-141-3p;hsa-miR-148a-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-589-3p;hsa-miR-590-3p | 12 | TGFB2 | Sponge network | -2.039 | 0 | -1.547 | 1.0E-5 | 0.272 |
| 55 | CASC2 |
hsa-miR-130b-3p;hsa-miR-142-3p;hsa-miR-148b-3p;hsa-miR-186-5p;hsa-miR-193b-3p;hsa-miR-21-5p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-454-3p;hsa-miR-590-3p;hsa-miR-590-5p | 11 | TGFB2 | Sponge network | -1.086 | 0 | -1.547 | 1.0E-5 | 0.266 |
| 56 | RP5-1198O20.4 | hsa-let-7g-5p;hsa-miR-195-3p;hsa-miR-195-5p;hsa-miR-204-5p;hsa-miR-26b-5p;hsa-miR-29b-2-5p;hsa-miR-30d-3p;hsa-miR-362-3p;hsa-miR-491-5p;hsa-miR-664a-3p;hsa-miR-664a-5p | 11 | HMGA2 | Sponge network | 0.401 | 0.05688 | 3.122 | 0.00011 | 0.265 |
| 57 | FAM225B | hsa-let-7a-5p;hsa-let-7g-5p;hsa-miR-26b-5p;hsa-miR-29b-2-5p;hsa-miR-3065-5p;hsa-miR-30b-5p;hsa-miR-30c-5p;hsa-miR-30d-5p;hsa-miR-664a-3p;hsa-miR-664a-5p | 10 | HMGA2 | Sponge network | 0.212 | 0.51382 | 3.122 | 0.00011 | 0.26 |
| 58 | FAM95B1 | hsa-miR-107;hsa-miR-182-5p;hsa-miR-193b-3p;hsa-miR-335-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-625-5p;hsa-miR-98-5p | 10 | TGFBR3 | Sponge network | -1.103 | 0.01335 | -3.002 | 0 | 0.259 |
| 59 | LINC00525 | hsa-let-7a-5p;hsa-let-7b-5p;hsa-let-7c-5p;hsa-let-7d-5p;hsa-miR-129-5p;hsa-miR-195-5p;hsa-miR-26b-5p;hsa-miR-29b-2-5p;hsa-miR-3065-5p;hsa-miR-30d-3p;hsa-miR-362-3p | 11 | HMGA2 | Sponge network | 3.158 | 0.00015 | 3.122 | 0.00011 | 0.257 |