This regulatory network was inferred from the input dataset. The miRNAs and mRNAs are
presented as round and rectangle nodes respectively. The numerical value popped up upon mouse over the gene node is the log2 transformed fold-change of the gene expression between the two groups. All of the nodes are clickable, and the detailed information of the miRNAs/mRNAs and related cancer pathway will be displayed in another window. The edges between nodes are supported by both interactions (predicted or experimentally verified) and correlations learnt from cancer dataset. The numerical value popped up upon mouse over the edge is the correlation beat value (effect size) between the two nodes. The experimental evidences of the edges reported in previous cancer studies are highlighted by red/orange color. All of these information can be accessed by the "mouse-over" action. This network shows a full map of the miRNA-mRNA regulation of the input gene list(s), and the hub miRNAs (with the high network degree/betweenness centrality) would be the potential cancer drivers or tumor suppressors. The full result table can be accessed in the "Regulations" tab.
"miRNACancerMAP" is also a network visualization tool for users to draw their regulatory network by personal customization. Users can set the complexity of the network by limiting the number of nodes or edges. And the color of the nodes can be defined by different categories of the mRNAs and miRNAs, such as Gene-Ontology, pathway, and expression status. Users can also select to use network degree or network betweenness centrality to define the node size. And edges can be black or colored by the correlation. Purple edge means negative correlation (mostly found between miRNA and mRNA), and blue edge means positive correlation (found in PPI or miRNA-miRNA sponge effect). We can also add the protein-protein interactions (PPI) into the network. This result will show the cluster of genes regulated by some specific miRNAs. Additionally, miRNA-miRNA edges can be added by the "miRNA sponge" button, presenting some clusters of miRNAs that have the interactions via sponge effect.
miRNA-gene regulations
| Num | microRNA | Gene | miRNA log2FC | miRNA pvalue | Gene log2FC | Gene pvalue | Interaction | Correlation beta | Correlation P-value | PMID | Reported in cancer studies |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | hsa-let-7a-5p | AIFM1 | -0.44 | 3.0E-5 | 0.03 | 0.68966 | TargetScan; miRNATAP | -0.1 | 0.00076 | NA | |
| 2 | hsa-let-7b-5p | AIFM1 | -0.25 | 0.02114 | 0.03 | 0.68966 | miRNATAP | -0.16 | 0 | NA | |
| 3 | hsa-let-7e-5p | AIFM1 | 0.09 | 0.51287 | 0.03 | 0.68966 | miRNATAP | -0.08 | 0.00189 | NA | |
| 4 | hsa-miR-125b-5p | AKT1 | -0.99 | 0 | 0.3 | 2.0E-5 | miRNAWalker2 validate; miRTarBase | -0.08 | 0.00021 | NA | |
| 5 | hsa-miR-26b-5p | AKT1 | -0.25 | 0.02852 | 0.3 | 2.0E-5 | miRNAWalker2 validate | -0.1 | 0.00023 | NA | |
| 6 | hsa-miR-183-5p | AKT2 | 1.43 | 0 | -0.27 | 1.0E-5 | mirMAP | -0.05 | 8.0E-5 | NA | |
| 7 | hsa-miR-18a-5p | AKT2 | 1.65 | 0 | -0.27 | 1.0E-5 | mirMAP | -0.05 | 0.00013 | NA | |
| 8 | hsa-miR-21-5p | AKT2 | 1.62 | 0 | -0.27 | 1.0E-5 | miRNAWalker2 validate | -0.07 | 0.00196 | NA | |
| 9 | hsa-miR-324-5p | AKT2 | 1.67 | 0 | -0.27 | 1.0E-5 | miRanda | -0.06 | 6.0E-5 | NA | |
| 10 | hsa-miR-339-5p | AKT2 | 1.08 | 0 | -0.27 | 1.0E-5 | mirMAP | -0.05 | 0.00043 | NA | |
| 11 | hsa-miR-708-5p | AKT2 | 1.54 | 0 | -0.27 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.07 | 0 | 22552290 | Together our findings suggest that reduced miR-708 expression leads to prostate cancer initiation progression and development by regulating the expression of CD44 as well as AKT2 miR-708 therefore may represent a novel therapeutic target or diagnostic and prognostic biomarker in prostate cancer |
| 12 | hsa-miR-766-3p | AKT2 | 1.07 | 0 | -0.27 | 1.0E-5 | mirMAP | -0.05 | 0.00735 | NA | |
| 13 | hsa-miR-139-5p | AKT3 | -1.46 | 0 | 0.76 | 5.0E-5 | PITA; miRanda | -0.2 | 7.0E-5 | NA | |
| 14 | hsa-miR-15a-5p | AKT3 | 0.78 | 0 | 0.76 | 5.0E-5 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.27 | 0.00011 | NA | |
| 15 | hsa-miR-17-3p | AKT3 | -0.01 | 0.90956 | 0.76 | 5.0E-5 | miRNATAP | -0.17 | 0.02246 | NA | |
| 16 | hsa-miR-17-5p | AKT3 | 1.42 | 0 | 0.76 | 5.0E-5 | TargetScan; miRNATAP | -0.21 | 1.0E-5 | NA | |
| 17 | hsa-miR-205-3p | AKT3 | 1.3 | 0 | 0.76 | 5.0E-5 | mirMAP | -0.25 | 0 | NA | |
| 18 | hsa-miR-20a-5p | AKT3 | 1.15 | 0 | 0.76 | 5.0E-5 | miRNATAP | -0.24 | 0 | NA | |
| 19 | hsa-miR-29b-3p | AKT3 | -0.11 | 0.51126 | 0.76 | 5.0E-5 | miRNATAP | -0.26 | 0 | 26512921 | MicroRNA 29B mir 29b regulates the Warburg effect in ovarian cancer by targeting AKT2 and AKT3 |
| 20 | hsa-miR-362-3p | AKT3 | -0.7 | 0 | 0.76 | 5.0E-5 | miRanda | -0.2 | 0.00031 | NA | |
| 21 | hsa-miR-362-5p | AKT3 | 0.41 | 0.04043 | 0.76 | 5.0E-5 | PITA; TargetScan; miRNATAP | -0.16 | 0.00012 | NA | |
| 22 | hsa-miR-501-3p | AKT3 | 0.82 | 0 | 0.76 | 5.0E-5 | miRNATAP | -0.12 | 0.03688 | NA | |
| 23 | hsa-miR-139-5p | APAF1 | -1.46 | 0 | 0.25 | 0.00843 | mirMAP | -0.07 | 0.01151 | NA | |
| 24 | hsa-miR-664a-3p | APAF1 | -0.26 | 0.05132 | 0.25 | 0.00843 | mirMAP | -0.13 | 3.0E-5 | NA | |
| 25 | hsa-miR-18a-5p | ATM | 1.65 | 0 | -0.09 | 0.45414 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.17 | 0 | 23437304; 25963391; 23857602; 23229340 | MicroRNA 18a attenuates DNA damage repair through suppressing the expression of ataxia telangiectasia mutated in colorectal cancer; Through in silico search the 3'UTR of Ataxia telangiectasia mutated ATM contains a conserved miR-18a binding site; Expression of ATM was down-regulated in CRC tumors p<0.0001 and inversely correlated with miR-18a expression r = -0.4562 p<0.01; This was further confirmed by the down-regulation of ATM protein by miR-18a; As ATM is a key enzyme in DNA damage repair we evaluated the effect of miR-18a on DNA double-strand breaks; miR-18a attenuates cellular repair of DNA double-strand breaks by directly suppressing ATM a key enzyme in DNA damage repair;However the upregulation of miR-18a suppressed the level of ataxia-telangiectasia mutated and attenuated DNA double-strand break repair after irradiation which re-sensitized the cervical cancer cells to radiotherapy by promoting apoptosis;Furthermore we used antisense oligonucleotides against micro RNAs miRNA or miRNA overexpression plasmids to study the role of miR-18a and -106a on ATM expression; Furthermore we identified that ERα activates miR-18a and -106a to downregulate ATM expression; We reveal a novel mechanism involving ERα and miR-18a and -106a regulation of ATM in breast cancer;MicroRNA 18a upregulates autophagy and ataxia telangiectasia mutated gene expression in HCT116 colon cancer cells; Previous studies showed that certain microRNAs including miR-18a potentially regulate ATM in cancer cells; However the mechanisms behind the modulation of ATM by miR-18a remain to be elucidated in colon cancer cells; In the present study we explored the impact of miR-18a on the autophagy process and ATM expression in HCT116 colon cancer cells; Western blotting and luciferase assays were implemented to explore the impact of miR-18a on ATM gene expression in HCT116 cells; Moreover miR-18a overexpression led to the upregulation of ATM expression and suppression of mTORC1 activity; Results of the present study pertaining to the role of miR-18a in regulating autophagy and ATM gene expression in colon cancer cells revealed a novel function for miR-18a in a critical cellular event and on a crucial gene with significant impacts in cancer development progression treatment and in other diseases |
| 26 | hsa-miR-19b-3p | ATM | 0.54 | 0.00062 | -0.09 | 0.45414 | miRNAWalker2 validate | -0.19 | 0 | NA | |
| 27 | hsa-miR-203a-3p | ATM | 0.04 | 0.88046 | -0.09 | 0.45414 | MirTarget | -0.1 | 0 | 24145123; 27542403 | miR 203 induces oxaliplatin resistance in colorectal cancer cells by negatively regulating ATM kinase; In silico analysis identified ataxia telangiectasia mutated ATM a primary mediator of the DNA damage response as a potential target of miR-203; Using TCGA database we identified a significant reverse correlation of miR-203 and ATM expression in CRC tissues; We validated ATM as a bona fide target of miR-203 in CRC cells; Mutation of the putative miR-203 binding site in the 3' untranslated region 3'UTR of the ATM mRNA abolished the inhibitory effect of miR-203 on ATM;MiR 203 inhibits tumor invasion and metastasis in gastric cancer by ATM; Our results showed that miR-203 was significantly downregulated in gastric cancer tissues and cells while ataxia telangiectasia mutated kinase ATM was upregulated in gastric cancer tissues and cells and was directly regulated by miR-203; ATM knockdown phenocopied the effect of miR-203 overexpression |
| 28 | hsa-miR-205-3p | ATM | 1.3 | 0 | -0.09 | 0.45414 | mirMAP | -0.09 | 0.00079 | NA | |
| 29 | hsa-miR-21-5p | ATM | 1.62 | 0 | -0.09 | 0.45414 | mirMAP | -0.14 | 0.00355 | 26289851 | MiR-21 is an oncomiR that is overexpressed in nearly all cancers including ATC; Hence suppression of miR-21 could pave the way for ATC therapy |
| 30 | hsa-miR-30b-5p | ATM | -0.4 | 0.00347 | -0.09 | 0.45414 | mirMAP | -0.1 | 0.01286 | 25202329 | However specific miRNAs are downregulated in ATC such as those of the miR-200 and miR-30 families which are important negative regulators of cell migration invasion and epithelial-to-mesenchymal transition EMT processes that are overactivated in ATC |
| 31 | hsa-miR-30c-5p | ATM | 0.02 | 0.88637 | -0.09 | 0.45414 | mirMAP | -0.12 | 0.00081 | NA | |
| 32 | hsa-miR-324-5p | ATM | 1.67 | 0 | -0.09 | 0.45414 | miRanda | -0.1 | 0.00178 | NA | |
| 33 | hsa-miR-339-5p | ATM | 1.08 | 0 | -0.09 | 0.45414 | miRanda | -0.14 | 0 | NA | |
| 34 | hsa-miR-455-5p | ATM | 1.41 | 0 | -0.09 | 0.45414 | miRanda | -0.22 | 0 | NA | |
| 35 | hsa-miR-500a-5p | ATM | 0.68 | 8.0E-5 | -0.09 | 0.45414 | mirMAP | -0.14 | 1.0E-5 | NA | |
| 36 | hsa-miR-576-5p | ATM | 0.94 | 0 | -0.09 | 0.45414 | mirMAP | -0.17 | 2.0E-5 | NA | |
| 37 | hsa-miR-590-5p | ATM | 0.76 | 0 | -0.09 | 0.45414 | mirMAP | -0.15 | 0.00038 | NA | |
| 38 | hsa-miR-92a-3p | ATM | 0.99 | 0 | -0.09 | 0.45414 | miRNAWalker2 validate | -0.1 | 0.01835 | NA | |
| 39 | hsa-miR-944 | ATM | 1.47 | 0 | -0.09 | 0.45414 | mirMAP | -0.09 | 0.00045 | NA | |
| 40 | hsa-miR-127-3p | BAD | -1.47 | 0 | 0.23 | 0.0411 | miRanda; miRNATAP | -0.11 | 1.0E-5 | NA | |
| 41 | hsa-miR-326 | BAD | -0.43 | 0.0415 | 0.23 | 0.0411 | miRanda | -0.16 | 0 | NA | |
| 42 | hsa-miR-17-5p | BCL2 | 1.42 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.29 | 0 | 25435430 | Combined overexpression of miR-16 and miR-17 greatly reduced Beclin-1 and Bcl-2 expressions respectively; miR-17 overexpression reduced cytoprotective autophagy by targeting Beclin-1 whereas overexpression of miR-16 potentiated paclitaxel induced apoptotic cell death by inhibiting anti-apoptotic protein Bcl-2 |
| 43 | hsa-miR-181b-5p | BCL2 | 0.97 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.37 | 0 | 25524579 | Moreover we also found miR-181 reduction was associated with increased Bcl-2 levels and miR-181 was further suggested to exert its pro-apoptotic function mainly through targeting Bcl-2 expression |
| 44 | hsa-miR-205-3p | BCL2 | 1.3 | 0 | -0.91 | 1.0E-5 | mirMAP | -0.23 | 0 | 23612742 | MicroRNA 205 a novel regulator of the anti apoptotic protein Bcl2 is downregulated in prostate cancer; In addition we describe the anti-apoptotic protein BCL2 as a target of miR-205 relevant for prostate cancer due to its role in prognosis of primary tumours and in the appearance of androgen independence; The repression of BCL2 by miR-205 was confirmed using reporter assays and western blotting; Consistent with its anti-apoptotic target BCL2 miR-205 promoted apoptosis in prostate cancer cells in response to DNA damage by cisplatin and doxorubicin in the prostate cancer cell lines PC3 and LnCap |
| 45 | hsa-miR-20a-3p | BCL2 | 1.35 | 0 | -0.91 | 1.0E-5 | mirMAP | -0.21 | 1.0E-5 | NA | |
| 46 | hsa-miR-20a-5p | BCL2 | 1.15 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.21 | 8.0E-5 | NA | |
| 47 | hsa-miR-21-5p | BCL2 | 1.62 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.44 | 0 | 21468550; 25994220; 25381586; 26555418; 23359184; 22964582; 21376256 | BCL-2 up-regulation could be achieved by miR-21 overexpression which prevented T24 cells from apoptosis induced by doxorubicin; Furthermore the miR-21 induced BCL-2 up-regulation could be cancelled by the PI3K inhibitor LY294002;Meanwhile miR-21 loss reduced STAT3 and Bcl-2 activation causing an increase in the apoptosis of tumour cells in CAC mice;Changes in the sensitivity of osteosarcoma cells to CDDP were examined after transfection with miR-21 mimics or anti-miR-21 or bcl-2 siRNA in combination with CDDP;The expression of Bax Bcl-2 and miR-21 in parental and paclitaxel-resistant cells was detected by RT-PCR and Western blotting;Resveratrol induces apoptosis of pancreatic cancers cells by inhibiting miR 21 regulation of BCL 2 expression; We also used Western blot to measure BCL-2 protein levels after down-regulation of miR-21 expression; Besides down-regulation of miR-21 expression can inhibit BCL-2 expression in PANC-1 CFPAC-1 and MIA Paca-2 cells; Over-expression of miR-21 expression can reverse down-regulation of BCL-2 expression and apoptosis induced by resveratrol; In this study we demonstrated that the effect of resveratrol on apoptosis is due to inhibiting miR-21 regulation of BCL-2 expression;Tumors harvested from these lungs have elevated levels of oncogenic miRNAs miR-21 and miR-155; are deficient for p53-regulated miRNAs; and have heightened expression of miR-34 target genes such as Met and Bcl-2;Bcl 2 upregulation induced by miR 21 via a direct interaction is associated with apoptosis and chemoresistance in MIA PaCa 2 pancreatic cancer cells; However the roles and mechanisms of miRNA miR-21 in regulation of Bcl-2 in pancreatic cancer remain to be elucidated; Then luciferase activity was observed after miR-21 mimics and pRL-TK plasmids containing wild-type and mutant 3'UTRs of Bcl-2 mRNA were co-transfected; Cells transfected with miR-21 inhibitor revealed an opposite trend. There was a significant increase in luciferase activity in the cells transfected with the wild-type pRL-TK plasmid in contrast to those transfected with the mutant one indicating that miR-21 promotes Bcl-2 expression by binding directly to the 3'UTR of Bcl-2 mRNA; Upregulation of Bcl-2 directly induced by miR-21 is associated with apoptosis chemoresistance and proliferation of MIA PaCa-2 pancreatic cancer cells |
| 48 | hsa-miR-224-3p | BCL2 | 0.5 | 0.00811 | -0.91 | 1.0E-5 | mirMAP | -0.15 | 0.00211 | 24796455 | In addition the expressions of Bcl2 mRNA and protein were 1.05 ± 0.04 and 0.21 ± 0.03 in the miR-224 ASO group significantly lower than that in the control group 4.87 ± 0.96 and 0.88 ± 0.09 P < 0.01 |
| 49 | hsa-miR-224-5p | BCL2 | 1.95 | 0 | -0.91 | 1.0E-5 | mirMAP | -0.35 | 0 | 24796455 | In addition the expressions of Bcl2 mRNA and protein were 1.05 ± 0.04 and 0.21 ± 0.03 in the miR-224 ASO group significantly lower than that in the control group 4.87 ± 0.96 and 0.88 ± 0.09 P < 0.01 |
| 50 | hsa-miR-24-2-5p | BCL2 | 1.27 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.49 | 0 | NA | |
| 51 | hsa-miR-29a-5p | BCL2 | 0.32 | 0.03704 | -0.91 | 1.0E-5 | mirMAP | -0.16 | 0.00917 | 20041405 | Subsequent investigation characterized two antiapoptotic molecules Bcl-2 and Mcl-1 as direct targets of miR-29; Furthermore silencing of Bcl-2 and Mcl-1 phenocopied the proapoptotic effect of miR-29 whereas overexpression of these proteins attenuated the effect of miR-29 |
| 52 | hsa-miR-365a-3p | BCL2 | 0.45 | 0.00234 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.62 | 0 | NA | |
| 53 | hsa-miR-450b-5p | BCL2 | 1.26 | 0 | -0.91 | 1.0E-5 | mirMAP | -0.3 | 0 | NA | |
| 54 | hsa-miR-451a | BCL2 | -1.24 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase | -0.09 | 0.00804 | 26516138; 23053883 | Western blot was used for the detection of the protein expression of Bcl-2 and Caspase 3 after the transfection of miR-451 mimics /inhibitors; Bioinformatics analysis demonstrated that Bcl-2 protein is a potential target gene for miR-451; Increased miR-451 expression may negatively regulate Bcl-2 mRNA and protein expression followed by affecting the protein expression of caspase 3 and accelerate the apoptosis in breast cancer indicating that miR-451 might influence the drug resistances of the Paclitaxel-resistant breast cancer cell line;The expression of miR-451 was analyzed by RT-PCR and the expressions of Bcl-2 AKT and phosphorylated AKT were analyzed by Western blotting; In comparison to the controls a significant increase in the expression of miR-451 was associated with significantly decreased expressions of Bcl-2 AKT and p-AKT and a significant increase in the apoptosis rate |
| 55 | hsa-miR-452-5p | BCL2 | 0.37 | 0.03485 | -0.91 | 1.0E-5 | mirMAP | -0.29 | 0 | NA | |
| 56 | hsa-miR-455-5p | BCL2 | 1.41 | 0 | -0.91 | 1.0E-5 | mirMAP | -0.67 | 0 | NA | |
| 57 | hsa-miR-542-3p | BCL2 | 0.64 | 0 | -0.91 | 1.0E-5 | mirMAP | -0.3 | 2.0E-5 | NA | |
| 58 | hsa-miR-590-5p | BCL2 | 0.76 | 0 | -0.91 | 1.0E-5 | miRanda | -0.21 | 0.00251 | NA | |
| 59 | hsa-miR-7-5p | BCL2 | 1.56 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; miRTarBase; mirMAP | -0.24 | 0 | 26464649; 25862909; 21750649 | Western blotting was used to evaluate the effect of miR-7 on Bcl2 in A549 and H460 cells; Moreover subsequent experiments showed that BCL-2 was downregulated by miR-7 at both transcriptional and translational levels; This study further extends the biological role of miR-7 in NSCLC A549 and H460 cells and identifies BCL-2 as a novel target possibly involved in miR-7-mediated growth suppression and apoptosis induction of NSCLC cells;miR-7 overexpression correlated with diminished BCL2 expression but there was no relationship between miR-7 and EGFR expression neither in tumour samples nor in the cell lines; Of the two postulated miR-7 target genes we examined BCL2 but not EGFR seems to be a possible miR-7 target in OC;Bioinformatics predictions revealed a potential binding site of miR-7 on 3'UTR of BCL-2 and it was further confirmed by luciferase assay; Moreover subsequent experiments showed that BCL-2 was downregulated by miR-7 at both transcriptional and translational levels; These results suggest that miR-7 regulates the expression of BCL-2 through direct 3'UTR interactions |
| 60 | hsa-miR-96-5p | BCL2 | 1.42 | 0 | -0.91 | 1.0E-5 | miRNAWalker2 validate; TargetScan | -0.11 | 0.02141 | NA | |
| 61 | hsa-let-7a-5p | BCL2L1 | -0.44 | 3.0E-5 | -0.15 | 0.08157 | TargetScan; miRNATAP | -0.09 | 0.01051 | 26915294; 20347499 | As a result transcript levels of the tumor-suppressive miR-15 and let-7 families increased which targeted and decreased the expression of the crucial prosurvival genes BCL-2 and BCL-XL respectively;The let 7 family of microRNAs inhibits Bcl xL expression and potentiates sorafenib induced apoptosis in human hepatocellular carcinoma; The effect of let-7 on Bcl-xL expression was examined by Western blot and a reporter assay; Microarray analysis followed by in silico target prediction identified let-7 microRNAs as being downregulated in Huh7 hepatoma cells in comparison with primary human hepatocytes as well as possessing a putative target site in the bcl-xl mRNA |
| 62 | hsa-let-7b-5p | BCL2L1 | -0.25 | 0.02114 | -0.15 | 0.08157 | miRNATAP | -0.09 | 0.00919 | 26915294; 20347499 | As a result transcript levels of the tumor-suppressive miR-15 and let-7 families increased which targeted and decreased the expression of the crucial prosurvival genes BCL-2 and BCL-XL respectively;The let 7 family of microRNAs inhibits Bcl xL expression and potentiates sorafenib induced apoptosis in human hepatocellular carcinoma; The effect of let-7 on Bcl-xL expression was examined by Western blot and a reporter assay; Microarray analysis followed by in silico target prediction identified let-7 microRNAs as being downregulated in Huh7 hepatoma cells in comparison with primary human hepatocytes as well as possessing a putative target site in the bcl-xl mRNA |
| 63 | hsa-let-7i-5p | BCL2L1 | 0.25 | 0.00427 | -0.15 | 0.08157 | miRNATAP | -0.11 | 0.01404 | NA | |
| 64 | hsa-miR-214-3p | BCL2L1 | -0.12 | 0.56496 | -0.15 | 0.08157 | mirMAP | -0.06 | 0.00064 | NA | |
| 65 | hsa-miR-342-5p | BCL2L1 | 0.86 | 0 | -0.15 | 0.08157 | miRNATAP | -0.08 | 0.00168 | NA | |
| 66 | hsa-miR-491-5p | BCL2L1 | -0.54 | 0.00136 | -0.15 | 0.08157 | miRNAWalker2 validate; miRTarBase; MirTarget; miRanda | -0.08 | 0.00064 | 25299770; 20039318; 23519249 | We found that miR-491-5p efficiently induces apoptosis in IGROV1-R10 cells by directly inhibiting BCL-XL expression and by inducing BIM accumulation in its dephosphorylated form;Functional screening identifies a microRNA miR 491 that induces apoptosis by targeting Bcl XL in colorectal cancer cells; We demonstrated that Bcl-XL was a direct target of miR-491 and its silencing contributed to miR-491-induced apoptosis; Our study provides a new regulation of Bcl-XL by miR-491 in colorectal cancer cells and suggests a therapeutic potential of miRNAs for treating colorectal cancer by targeting Bcl-XL;Targeted site prediction indicated that both Bcl-XL and TP53 contain miR-491-5p recognizing sites in their 3' UTRs; Overexpression of miR-491-5p in the pancreatic cancer cell line SW1990 effectively inhibited both endogenous Bcl-XL and TP53 gene expressions |
| 67 | hsa-miR-625-5p | BCL2L1 | 0.35 | 0.02861 | -0.15 | 0.08157 | MirTarget | -0.09 | 0.00015 | NA | |
| 68 | hsa-miR-338-5p | BID | -1.72 | 0 | 0.97 | 0 | MirTarget | -0.15 | 0 | NA | |
| 69 | hsa-miR-511-5p | BID | 0.21 | 0.23237 | 0.97 | 0 | MirTarget | -0.07 | 0.02908 | NA | |
| 70 | hsa-miR-29c-3p | BIRC2 | -2.08 | 0 | 0.18 | 0.17933 | MirTarget | -0.1 | 0.00021 | NA | |
| 71 | hsa-miR-624-5p | BIRC2 | -0.23 | 0.11049 | 0.18 | 0.17933 | MirTarget | -0.09 | 0.03805 | NA | |
| 72 | hsa-miR-369-3p | BIRC3 | -1.23 | 0 | 0.2 | 0.3995 | MirTarget; miRNATAP | -0.25 | 0 | NA | |
| 73 | hsa-miR-34a-5p | CAPN1 | -0.12 | 0.364 | -0.35 | 0.00055 | miRNAWalker2 validate | -0.18 | 0 | NA | |
| 74 | hsa-miR-491-5p | CAPN1 | -0.54 | 0.00136 | -0.35 | 0.00055 | miRanda | -0.12 | 2.0E-5 | NA | |
| 75 | hsa-miR-1468-5p | CAPN2 | -1.12 | 0 | -0.39 | 3.0E-5 | MirTarget | -0.06 | 0.00726 | NA | |
| 76 | hsa-miR-16-2-3p | CAPN2 | 1.16 | 0 | -0.39 | 3.0E-5 | mirMAP | -0.1 | 7.0E-5 | NA | |
| 77 | hsa-miR-20a-3p | CAPN2 | 1.35 | 0 | -0.39 | 3.0E-5 | MirTarget | -0.08 | 7.0E-5 | NA | |
| 78 | hsa-miR-320a | CAPN2 | 0.8 | 0 | -0.39 | 3.0E-5 | miRanda | -0.14 | 0 | NA | |
| 79 | hsa-miR-320b | CAPN2 | 1.23 | 0 | -0.39 | 3.0E-5 | miRanda | -0.11 | 0 | NA | |
| 80 | hsa-miR-421 | CAPN2 | 0.95 | 0 | -0.39 | 3.0E-5 | miRanda | -0.05 | 0.03514 | NA | |
| 81 | hsa-miR-590-5p | CAPN2 | 0.76 | 0 | -0.39 | 3.0E-5 | miRanda | -0.1 | 0.00115 | NA | |
| 82 | hsa-miR-125a-5p | CASP10 | 0.22 | 0.11955 | 0.04 | 0.78895 | mirMAP | -0.16 | 0.00042 | NA | |
| 83 | hsa-miR-125b-5p | CASP10 | -0.99 | 0 | 0.04 | 0.78895 | mirMAP | -0.09 | 0.04635 | NA | |
| 84 | hsa-miR-129-5p | CASP10 | -0.25 | 0.43645 | 0.04 | 0.78895 | mirMAP | -0.08 | 5.0E-5 | NA | |
| 85 | hsa-miR-145-3p | CASP10 | -1.31 | 0 | 0.04 | 0.78895 | mirMAP | -0.19 | 5.0E-5 | NA | |
| 86 | hsa-miR-145-5p | CASP10 | -0.47 | 0.00069 | 0.04 | 0.78895 | mirMAP | -0.21 | 0 | NA | |
| 87 | hsa-miR-19b-3p | CASP10 | 0.54 | 0.00062 | 0.04 | 0.78895 | MirTarget; mirMAP | -0.09 | 0.02915 | NA | |
| 88 | hsa-miR-217 | CASP10 | -1.35 | 0 | 0.04 | 0.78895 | mirMAP | -0.06 | 0.02452 | NA | |
| 89 | hsa-miR-28-3p | CASP10 | 0.46 | 0 | 0.04 | 0.78895 | mirMAP | -0.23 | 0.00093 | NA | |
| 90 | hsa-miR-320a | CASP10 | 0.8 | 0 | 0.04 | 0.78895 | miRanda | -0.13 | 0.00592 | NA | |
| 91 | hsa-miR-320b | CASP10 | 1.23 | 0 | 0.04 | 0.78895 | miRanda | -0.09 | 0.01731 | NA | |
| 92 | hsa-miR-339-5p | CASP10 | 1.08 | 0 | 0.04 | 0.78895 | MirTarget | -0.1 | 0.00304 | NA | |
| 93 | hsa-miR-361-5p | CASP10 | 0.08 | 0.31404 | 0.04 | 0.78895 | miRanda | -0.23 | 0.00452 | NA | |
| 94 | hsa-miR-589-3p | CASP10 | 1.6 | 0 | 0.04 | 0.78895 | mirMAP | -0.06 | 0.0405 | NA | |
| 95 | hsa-miR-744-3p | CASP10 | 1.19 | 0 | 0.04 | 0.78895 | mirMAP | -0.15 | 1.0E-5 | NA | |
| 96 | hsa-let-7e-5p | CASP3 | 0.09 | 0.51287 | 0.16 | 0.06548 | MirTarget; miRNATAP | -0.06 | 0.03025 | NA | |
| 97 | hsa-miR-193b-5p | CASP3 | 1.1 | 0 | 0.16 | 0.06548 | MirTarget | -0.1 | 1.0E-5 | NA | |
| 98 | hsa-miR-382-5p | CASP3 | -0.51 | 0.00312 | 0.16 | 0.06548 | MirTarget | -0.07 | 0.00132 | NA | |
| 99 | hsa-miR-106b-5p | CASP7 | 0.5 | 1.0E-5 | 0.36 | 0.00041 | miRNAWalker2 validate | -0.1 | 0.01045 | 22986525 | MicroRNA 106b 25 cluster expression is associated with early disease recurrence and targets caspase 7 and focal adhesion in human prostate cancer; Moreover increased tumor miR-106b expression was associated with disease recurrence and the combination of high miR-106b and low CASP7 caspase-7 expressions in primary tumors was an independent predictor of early disease recurrence adjusted hazard ratio=4.1; 95% confidence interval: 1.6-12.3; The approach revealed that CASP7 is a direct target of miR-106b which was confirmed by western blot analysis and a 3'-untranslated region reporter assay; Moreover selected phenotypes induced by miR-106b knockdown in DU145 human prostate cancer cells did not develop when both miR-106b and CASP7 expression were inhibited |
| 100 | hsa-miR-23b-3p | CASP7 | -0.5 | 3.0E-5 | 0.36 | 0.00041 | MirTarget | -0.09 | 0.01326 | NA | |
| 101 | hsa-miR-26b-5p | CASP7 | -0.25 | 0.02852 | 0.36 | 0.00041 | miRNAWalker2 validate | -0.08 | 0.04203 | NA | |
| 102 | hsa-miR-3065-3p | CASP7 | -0.54 | 0.02132 | 0.36 | 0.00041 | MirTarget | -0.05 | 0.00424 | NA | |
| 103 | hsa-miR-335-5p | CASP7 | -1.17 | 0 | 0.36 | 0.00041 | MirTarget | -0.09 | 0.0001 | NA | |
| 104 | hsa-miR-361-5p | CASP7 | 0.08 | 0.31404 | 0.36 | 0.00041 | PITA; miRanda | -0.33 | 0 | NA | |
| 105 | hsa-miR-664a-3p | CASP7 | -0.26 | 0.05132 | 0.36 | 0.00041 | MirTarget | -0.14 | 3.0E-5 | NA | |
| 106 | hsa-miR-143-3p | CASP8 | -1.34 | 0 | 0.61 | 0 | MirTarget | -0.05 | 0.02691 | NA | |
| 107 | hsa-miR-217 | CASP8 | -1.35 | 0 | 0.61 | 0 | miRanda | -0.08 | 7.0E-5 | NA | |
| 108 | hsa-miR-17-5p | CASP9 | 1.42 | 0 | -0.46 | 0 | TargetScan | -0.07 | 0.00893 | NA | |
| 109 | hsa-miR-193b-3p | CASP9 | 1.73 | 0 | -0.46 | 0 | miRNAWalker2 validate | -0.15 | 0 | NA | |
| 110 | hsa-miR-424-5p | CASP9 | 1.48 | 0 | -0.46 | 0 | mirMAP | -0.14 | 0 | NA | |
| 111 | hsa-miR-450b-5p | CASP9 | 1.26 | 0 | -0.46 | 0 | mirMAP | -0.1 | 0.00019 | NA | |
| 112 | hsa-miR-7-5p | CASP9 | 1.56 | 0 | -0.46 | 0 | miRNAWalker2 validate | -0.1 | 0 | NA | |
| 113 | hsa-miR-103a-2-5p | CFLAR | 1.38 | 0 | -0.35 | 0.00043 | mirMAP | -0.06 | 0.0017 | NA | |
| 114 | hsa-miR-125a-5p | CFLAR | 0.22 | 0.11955 | -0.35 | 0.00043 | miRanda | -0.08 | 0.01365 | NA | |
| 115 | hsa-miR-130a-3p | CFLAR | 0.67 | 0 | -0.35 | 0.00043 | mirMAP | -0.13 | 1.0E-5 | NA | |
| 116 | hsa-miR-130b-3p | CFLAR | 1.45 | 0 | -0.35 | 0.00043 | mirMAP | -0.18 | 0 | NA | |
| 117 | hsa-miR-15b-5p | CFLAR | 0.85 | 0 | -0.35 | 0.00043 | mirMAP | -0.1 | 0.00441 | NA | |
| 118 | hsa-miR-199a-5p | CFLAR | -0.36 | 0.04351 | -0.35 | 0.00043 | mirMAP | -0.09 | 0.00029 | NA | |
| 119 | hsa-miR-199b-5p | CFLAR | -1.08 | 0 | -0.35 | 0.00043 | mirMAP | -0.05 | 0.02888 | NA | |
| 120 | hsa-miR-301a-3p | CFLAR | 2.14 | 0 | -0.35 | 0.00043 | mirMAP | -0.07 | 0.00024 | NA | |
| 121 | hsa-miR-320a | CFLAR | 0.8 | 0 | -0.35 | 0.00043 | miRanda | -0.08 | 0.01023 | NA | |
| 122 | hsa-miR-320b | CFLAR | 1.23 | 0 | -0.35 | 0.00043 | miRanda | -0.09 | 0.00049 | NA | |
| 123 | hsa-miR-339-5p | CFLAR | 1.08 | 0 | -0.35 | 0.00043 | miRanda | -0.08 | 0.00074 | NA | |
| 124 | hsa-miR-34c-5p | CFLAR | 0.57 | 0.02241 | -0.35 | 0.00043 | miRanda | -0.07 | 0.0001 | NA | |
| 125 | hsa-miR-374b-5p | CFLAR | -0.29 | 0.00415 | -0.35 | 0.00043 | MirTarget | -0.09 | 0.04376 | NA | |
| 126 | hsa-miR-421 | CFLAR | 0.95 | 0 | -0.35 | 0.00043 | miRanda | -0.1 | 0.00039 | NA | |
| 127 | hsa-miR-454-3p | CFLAR | 1.54 | 0 | -0.35 | 0.00043 | mirMAP | -0.05 | 0.03419 | NA | |
| 128 | hsa-miR-550a-5p | CFLAR | 1.49 | 0 | -0.35 | 0.00043 | mirMAP | -0.06 | 0.01266 | NA | |
| 129 | hsa-miR-708-5p | CFLAR | 1.54 | 0 | -0.35 | 0.00043 | mirMAP | -0.12 | 0 | NA | |
| 130 | hsa-miR-582-5p | CHP2 | -0.04 | 0.82183 | 0.03 | 0.94573 | miRNATAP | -0.3 | 0.01868 | NA | |
| 131 | hsa-miR-15a-5p | CHUK | 0.78 | 0 | 0.05 | 0.47292 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.1 | 0.0001 | NA | |
| 132 | hsa-miR-342-3p | CHUK | 1 | 0 | 0.05 | 0.47292 | miRanda | -0.06 | 0.00055 | NA | |
| 133 | hsa-miR-125a-5p | CSF2RB | 0.22 | 0.11955 | -0.37 | 0.08597 | miRanda | -0.14 | 0.04425 | NA | |
| 134 | hsa-miR-19a-3p | CSF2RB | 1.36 | 0 | -0.37 | 0.08597 | MirTarget | -0.32 | 0 | NA | |
| 135 | hsa-miR-19b-3p | CSF2RB | 0.54 | 0.00062 | -0.37 | 0.08597 | MirTarget | -0.36 | 0 | NA | |
| 136 | hsa-miR-452-3p | CSF2RB | 1.1 | 0 | -0.37 | 0.08597 | mirMAP | -0.2 | 1.0E-5 | NA | |
| 137 | hsa-miR-455-5p | CSF2RB | 1.41 | 0 | -0.37 | 0.08597 | miRanda | -0.35 | 0 | NA | |
| 138 | hsa-miR-485-3p | CSF2RB | 0.36 | 0.09294 | -0.37 | 0.08597 | MirTarget | -0.12 | 0.0086 | NA | |
| 139 | hsa-miR-493-3p | CSF2RB | -0.59 | 0.00436 | -0.37 | 0.08597 | MirTarget | -0.12 | 0.00918 | NA | |
| 140 | hsa-miR-576-5p | CSF2RB | 0.94 | 0 | -0.37 | 0.08597 | miRNATAP | -0.19 | 0.00484 | NA | |
| 141 | hsa-miR-125a-3p | CYCS | 0.12 | 0.44483 | -0.13 | 0.13395 | miRanda | -0.11 | 3.0E-5 | NA | |
| 142 | hsa-miR-127-3p | DFFA | -1.47 | 0 | 0.11 | 0.16666 | miRanda | -0.07 | 1.0E-5 | NA | |
| 143 | hsa-miR-129-5p | DFFA | -0.25 | 0.43645 | 0.11 | 0.16666 | miRanda; mirMAP | -0.05 | 0 | NA | |
| 144 | hsa-miR-145-5p | DFFA | -0.47 | 0.00069 | 0.11 | 0.16666 | miRNAWalker2 validate; miRTarBase | -0.09 | 0.00016 | NA | |
| 145 | hsa-miR-148a-3p | DFFA | -0.43 | 0.00954 | 0.11 | 0.16666 | mirMAP | -0.09 | 0 | NA | |
| 146 | hsa-miR-26a-2-3p | DFFA | -0.44 | 0.00326 | 0.11 | 0.16666 | mirMAP | -0.05 | 0.01279 | NA | |
| 147 | hsa-miR-26a-5p | DFFA | -1.09 | 0 | 0.11 | 0.16666 | mirMAP | -0.17 | 0 | NA | |
| 148 | hsa-miR-30a-3p | DFFA | -2.26 | 0 | 0.11 | 0.16666 | mirMAP | -0.06 | 9.0E-5 | NA | |
| 149 | hsa-miR-30e-3p | DFFA | -0.89 | 0 | 0.11 | 0.16666 | mirMAP | -0.08 | 0.00477 | NA | |
| 150 | hsa-miR-374a-5p | DFFA | -0.62 | 0 | 0.11 | 0.16666 | mirMAP | -0.08 | 0.03837 | NA |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | POSITIVE REGULATION OF RESPONSE TO STIMULUS | 54 | 1929 | 6.125e-39 | 2.85e-35 |
| 2 | INTRACELLULAR SIGNAL TRANSDUCTION | 50 | 1572 | 7.745e-38 | 1.802e-34 |
| 3 | CELL DEATH | 41 | 1001 | 5.008e-34 | 5.826e-31 |
| 4 | EXTRINSIC APOPTOTIC SIGNALING PATHWAY | 22 | 99 | 4.667e-34 | 5.826e-31 |
| 5 | REGULATION OF CELL DEATH | 46 | 1472 | 1.007e-33 | 9.376e-31 |
| 6 | POSITIVE REGULATION OF PROTEIN METABOLIC PROCESS | 46 | 1492 | 1.837e-33 | 1.425e-30 |
| 7 | APOPTOTIC SIGNALING PATHWAY | 28 | 289 | 8.68e-33 | 5.77e-30 |
| 8 | CELLULAR RESPONSE TO ORGANIC SUBSTANCE | 48 | 1848 | 7.665e-32 | 4.458e-29 |
| 9 | POSITIVE REGULATION OF CELL COMMUNICATION | 45 | 1532 | 1.175e-31 | 6.076e-29 |
| 10 | REGULATION OF INTRACELLULAR SIGNAL TRANSDUCTION | 46 | 1656 | 1.872e-31 | 8.711e-29 |
| 11 | POSITIVE REGULATION OF MOLECULAR FUNCTION | 47 | 1791 | 3.364e-31 | 1.423e-28 |
| 12 | POSITIVE REGULATION OF INTRACELLULAR SIGNAL TRANSDUCTION | 37 | 876 | 9.652e-31 | 3.742e-28 |
| 13 | EXTRINSIC APOPTOTIC SIGNALING PATHWAY VIA DEATH DOMAIN RECEPTORS | 16 | 39 | 7.65e-30 | 2.738e-27 |
| 14 | POSITIVE REGULATION OF I KAPPAB KINASE NF KAPPAB SIGNALING | 23 | 179 | 1.014e-29 | 3.37e-27 |
| 15 | REGULATION OF CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY | 24 | 213 | 1.328e-29 | 4.119e-27 |
| 16 | NEGATIVE REGULATION OF CELL DEATH | 36 | 872 | 1.823e-29 | 5.303e-27 |
| 17 | REGULATION OF I KAPPAB KINASE NF KAPPAB SIGNALING | 24 | 233 | 1.229e-28 | 3.364e-26 |
| 18 | ACTIVATION OF CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY | 19 | 95 | 1.849e-28 | 4.529e-26 |
| 19 | RESPONSE TO OXYGEN CONTAINING COMPOUND | 41 | 1381 | 1.77e-28 | 4.529e-26 |
| 20 | RESPONSE TO CYTOKINE | 33 | 714 | 2.4e-28 | 5.583e-26 |
| 21 | IMMUNE SYSTEM PROCESS | 46 | 1984 | 5.176e-28 | 1.147e-25 |
| 22 | ZYMOGEN ACTIVATION | 19 | 112 | 5.51e-27 | 1.165e-24 |
| 23 | POSITIVE REGULATION OF CATALYTIC ACTIVITY | 40 | 1518 | 1.076e-25 | 2.178e-23 |
| 24 | NEGATIVE REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY | 17 | 98 | 2.303e-24 | 4.465e-22 |
| 25 | POSITIVE REGULATION OF PEPTIDASE ACTIVITY | 19 | 154 | 3.32e-24 | 6.178e-22 |
| 26 | POSITIVE REGULATION OF CELL DEATH | 28 | 605 | 8.365e-24 | 1.497e-21 |
| 27 | REGULATION OF IMMUNE SYSTEM PROCESS | 37 | 1403 | 1.753e-23 | 3.021e-21 |
| 28 | REGULATION OF PEPTIDASE ACTIVITY | 24 | 392 | 3.666e-23 | 6.092e-21 |
| 29 | PROTEIN MATURATION | 21 | 265 | 1.342e-22 | 2.154e-20 |
| 30 | REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY | 18 | 153 | 1.453e-22 | 2.254e-20 |
| 31 | CELLULAR RESPONSE TO CYTOKINE STIMULUS | 27 | 606 | 1.747e-22 | 2.622e-20 |
| 32 | ACTIVATION OF IMMUNE RESPONSE | 24 | 427 | 2.783e-22 | 4.046e-20 |
| 33 | REGULATION OF IMMUNE RESPONSE | 30 | 858 | 4.458e-22 | 6.285e-20 |
| 34 | POSITIVE REGULATION OF IMMUNE RESPONSE | 26 | 563 | 5.152e-22 | 7.051e-20 |
| 35 | REGULATION OF PROTEOLYSIS | 28 | 711 | 6.634e-22 | 8.819e-20 |
| 36 | REGULATION OF APOPTOTIC SIGNALING PATHWAY | 22 | 363 | 4.06e-21 | 5.248e-19 |
| 37 | POSITIVE REGULATION OF DEFENSE RESPONSE | 22 | 364 | 4.31e-21 | 5.421e-19 |
| 38 | RESPONSE TO NITROGEN COMPOUND | 29 | 859 | 7.096e-21 | 8.689e-19 |
| 39 | POSITIVE REGULATION OF IMMUNE SYSTEM PROCESS | 29 | 867 | 9.159e-21 | 1.093e-18 |
| 40 | CYTOKINE MEDIATED SIGNALING PATHWAY | 23 | 452 | 2.283e-20 | 2.655e-18 |
| 41 | ACTIVATION OF INNATE IMMUNE RESPONSE | 18 | 204 | 2.941e-20 | 3.337e-18 |
| 42 | ACTIVATION OF CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 9 | 13 | 5.546e-20 | 6.144e-18 |
| 43 | PATTERN RECOGNITION RECEPTOR SIGNALING PATHWAY | 15 | 109 | 5.724e-20 | 6.194e-18 |
| 44 | RESPONSE TO ABIOTIC STIMULUS | 30 | 1024 | 6.75e-20 | 7.138e-18 |
| 45 | TOLL LIKE RECEPTOR SIGNALING PATHWAY | 14 | 85 | 7.586e-20 | 7.844e-18 |
| 46 | POSITIVE REGULATION OF PROTEOLYSIS | 21 | 363 | 9.678e-20 | 9.79e-18 |
| 47 | POSITIVE REGULATION OF PROTEIN MODIFICATION PROCESS | 31 | 1135 | 9.98e-20 | 9.88e-18 |
| 48 | RESPONSE TO MOLECULE OF BACTERIAL ORIGIN | 20 | 321 | 1.915e-19 | 1.857e-17 |
| 49 | CELLULAR RESPONSE TO OXYGEN CONTAINING COMPOUND | 27 | 799 | 2.24e-19 | 2.127e-17 |
| 50 | RESPONSE TO BIOTIC STIMULUS | 28 | 886 | 2.338e-19 | 2.176e-17 |
| 51 | REGULATION OF PHOSPHORUS METABOLIC PROCESS | 35 | 1618 | 2.936e-19 | 2.679e-17 |
| 52 | NEGATIVE REGULATION OF APOPTOTIC SIGNALING PATHWAY | 17 | 200 | 6.994e-19 | 6.259e-17 |
| 53 | POSITIVE REGULATION OF INNATE IMMUNE RESPONSE | 18 | 246 | 8.745e-19 | 7.677e-17 |
| 54 | RESPONSE TO ENDOGENOUS STIMULUS | 33 | 1450 | 1.07e-18 | 9.222e-17 |
| 55 | POSITIVE REGULATION OF PHOSPHATE METABOLIC PROCESS | 29 | 1036 | 1.179e-18 | 9.795e-17 |
| 56 | POSITIVE REGULATION OF PHOSPHORUS METABOLIC PROCESS | 29 | 1036 | 1.179e-18 | 9.795e-17 |
| 57 | REGULATION OF TRANSFERASE ACTIVITY | 28 | 946 | 1.31e-18 | 1.069e-16 |
| 58 | REGULATION OF KINASE ACTIVITY | 26 | 776 | 1.532e-18 | 1.226e-16 |
| 59 | REGULATION OF RESPONSE TO STRESS | 33 | 1468 | 1.554e-18 | 1.226e-16 |
| 60 | POSITIVE REGULATION OF APOPTOTIC SIGNALING PATHWAY | 16 | 171 | 1.749e-18 | 1.356e-16 |
| 61 | POSITIVE REGULATION OF CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 9 | 17 | 1.864e-18 | 1.421e-16 |
| 62 | RESPONSE TO TUMOR NECROSIS FACTOR | 17 | 233 | 9.493e-18 | 7.125e-16 |
| 63 | PHOSPHORYLATION | 30 | 1228 | 1.067e-17 | 7.883e-16 |
| 64 | REGULATION OF PROTEIN MODIFICATION PROCESS | 34 | 1710 | 1.646e-17 | 1.197e-15 |
| 65 | I KAPPAB KINASE NF KAPPAB SIGNALING | 12 | 70 | 2.319e-17 | 1.66e-15 |
| 66 | NEGATIVE REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY VIA DEATH DOMAIN RECEPTORS | 10 | 34 | 3.12e-17 | 2.199e-15 |
| 67 | POSITIVE REGULATION OF KINASE ACTIVITY | 21 | 482 | 3.181e-17 | 2.209e-15 |
| 68 | REGULATION OF INNATE IMMUNE RESPONSE | 19 | 357 | 3.288e-17 | 2.25e-15 |
| 69 | REGULATION OF CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 9 | 22 | 3.758e-17 | 2.534e-15 |
| 70 | POSITIVE REGULATION OF NF KAPPAB TRANSCRIPTION FACTOR ACTIVITY | 14 | 132 | 4.79e-17 | 3.184e-15 |
| 71 | CELLULAR RESPONSE TO ABIOTIC STIMULUS | 17 | 263 | 7.352e-17 | 4.818e-15 |
| 72 | CELLULAR RESPONSE TO NITROGEN COMPOUND | 21 | 505 | 8.143e-17 | 5.262e-15 |
| 73 | RESPONSE TO EXTERNAL STIMULUS | 34 | 1821 | 1.127e-16 | 7.182e-15 |
| 74 | CELLULAR RESPONSE TO MECHANICAL STIMULUS | 12 | 80 | 1.276e-16 | 8.024e-15 |
| 75 | REGULATION OF DEFENSE RESPONSE | 24 | 759 | 1.624e-16 | 1.008e-14 |
| 76 | REGULATION OF RESPONSE TO CYTOKINE STIMULUS | 14 | 144 | 1.665e-16 | 1.019e-14 |
| 77 | PROTEIN PHOSPHORYLATION | 26 | 944 | 1.801e-16 | 1.074e-14 |
| 78 | POSITIVE REGULATION OF SEQUENCE SPECIFIC DNA BINDING TRANSCRIPTION FACTOR ACTIVITY | 16 | 228 | 1.795e-16 | 1.074e-14 |
| 79 | RESPONSE TO BACTERIUM | 21 | 528 | 1.991e-16 | 1.158e-14 |
| 80 | ACTIVATION OF PROTEIN KINASE ACTIVITY | 17 | 279 | 1.981e-16 | 1.158e-14 |
| 81 | REGULATION OF HYDROLASE ACTIVITY | 29 | 1327 | 8.637e-16 | 4.961e-14 |
| 82 | PHOSPHATE CONTAINING COMPOUND METABOLIC PROCESS | 34 | 1977 | 1.354e-15 | 7.683e-14 |
| 83 | IMMUNE RESPONSE REGULATING CELL SURFACE RECEPTOR SIGNALING PATHWAY | 17 | 323 | 2.27e-15 | 1.272e-13 |
| 84 | REGULATION OF TUMOR NECROSIS FACTOR MEDIATED SIGNALING PATHWAY | 10 | 50 | 2.332e-15 | 1.277e-13 |
| 85 | REGULATION OF CELL PROLIFERATION | 30 | 1496 | 2.31e-15 | 1.277e-13 |
| 86 | INFLAMMATORY RESPONSE | 19 | 454 | 2.717e-15 | 1.47e-13 |
| 87 | SIGNAL TRANSDUCTION IN ABSENCE OF LIGAND | 9 | 33 | 2.821e-15 | 1.492e-13 |
| 88 | EXTRINSIC APOPTOTIC SIGNALING PATHWAY IN ABSENCE OF LIGAND | 9 | 33 | 2.821e-15 | 1.492e-13 |
| 89 | POSITIVE REGULATION OF TRANSFERASE ACTIVITY | 21 | 616 | 4.277e-15 | 2.236e-13 |
| 90 | FC EPSILON RECEPTOR SIGNALING PATHWAY | 13 | 142 | 4.815e-15 | 2.489e-13 |
| 91 | NEURON APOPTOTIC PROCESS | 9 | 35 | 5.134e-15 | 2.597e-13 |
| 92 | REGULATION OF PROTEIN SERINE THREONINE KINASE ACTIVITY | 19 | 470 | 5.094e-15 | 2.597e-13 |
| 93 | RESPONSE TO LIPID | 24 | 888 | 5.376e-15 | 2.69e-13 |
| 94 | REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY VIA DEATH DOMAIN RECEPTORS | 10 | 55 | 6.545e-15 | 3.24e-13 |
| 95 | PROTEOLYSIS | 27 | 1208 | 6.935e-15 | 3.397e-13 |
| 96 | CELLULAR RESPONSE TO STRESS | 30 | 1565 | 7.728e-15 | 3.746e-13 |
| 97 | POSITIVE REGULATION OF HYDROLASE ACTIVITY | 24 | 905 | 8.17e-15 | 3.919e-13 |
| 98 | INTRINSIC APOPTOTIC SIGNALING PATHWAY | 13 | 152 | 1.178e-14 | 5.593e-13 |
| 99 | TUMOR NECROSIS FACTOR MEDIATED SIGNALING PATHWAY | 12 | 118 | 1.62e-14 | 7.615e-13 |
| 100 | RESPONSE TO MECHANICAL STIMULUS | 14 | 210 | 3.354e-14 | 1.561e-12 |
| 101 | CELLULAR RESPONSE TO EXTERNAL STIMULUS | 15 | 264 | 3.934e-14 | 1.813e-12 |
| 102 | RESPONSE TO HORMONE | 23 | 893 | 6.313e-14 | 2.88e-12 |
| 103 | NUCLEOTIDE BINDING DOMAIN LEUCINE RICH REPEAT CONTAINING RECEPTOR SIGNALING PATHWAY | 8 | 28 | 6.966e-14 | 3.147e-12 |
| 104 | NEURON DEATH | 9 | 47 | 9.566e-14 | 4.28e-12 |
| 105 | MYD88 INDEPENDENT TOLL LIKE RECEPTOR SIGNALING PATHWAY | 8 | 30 | 1.304e-13 | 5.779e-12 |
| 106 | POSITIVE REGULATION OF CELLULAR PROTEIN LOCALIZATION | 16 | 360 | 2.273e-13 | 9.976e-12 |
| 107 | REGULATION OF SEQUENCE SPECIFIC DNA BINDING TRANSCRIPTION FACTOR ACTIVITY | 16 | 365 | 2.808e-13 | 1.221e-11 |
| 108 | CYTOPLASMIC PATTERN RECOGNITION RECEPTOR SIGNALING PATHWAY | 8 | 33 | 3.067e-13 | 1.321e-11 |
| 109 | REGULATION OF NEURON DEATH | 14 | 252 | 4.123e-13 | 1.76e-11 |
| 110 | IMMUNE RESPONSE | 24 | 1100 | 5.716e-13 | 2.418e-11 |
| 111 | FC RECEPTOR SIGNALING PATHWAY | 13 | 206 | 6.052e-13 | 2.537e-11 |
| 112 | CELLULAR RESPONSE TO ENDOGENOUS STIMULUS | 23 | 1008 | 7.89e-13 | 3.278e-11 |
| 113 | POSITIVE REGULATION OF GENE EXPRESSION | 29 | 1733 | 8.261e-13 | 3.402e-11 |
| 114 | RESPONSE TO ORGANIC CYCLIC COMPOUND | 22 | 917 | 1.043e-12 | 4.258e-11 |
| 115 | REGULATION OF CELLULAR PROTEIN LOCALIZATION | 18 | 552 | 1.138e-12 | 4.605e-11 |
| 116 | RESPONSE TO PEPTIDE | 16 | 404 | 1.321e-12 | 5.3e-11 |
| 117 | POSITIVE REGULATION OF ESTABLISHMENT OF PROTEIN LOCALIZATION | 17 | 514 | 4.251e-12 | 1.691e-10 |
| 118 | REGULATION OF NECROTIC CELL DEATH | 7 | 26 | 4.413e-12 | 1.74e-10 |
| 119 | INTRINSIC APOPTOTIC SIGNALING PATHWAY IN RESPONSE TO DNA DAMAGE | 9 | 71 | 4.873e-12 | 1.905e-10 |
| 120 | HOMEOSTATIC PROCESS | 25 | 1337 | 4.95e-12 | 1.919e-10 |
| 121 | POSITIVE REGULATION OF INTRACELLULAR TRANSPORT | 15 | 370 | 5.2e-12 | 1.999e-10 |
| 122 | REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY IN ABSENCE OF LIGAND | 8 | 46 | 5.547e-12 | 2.116e-10 |
| 123 | T CELL RECEPTOR SIGNALING PATHWAY | 11 | 146 | 6.044e-12 | 2.286e-10 |
| 124 | ANTIGEN RECEPTOR MEDIATED SIGNALING PATHWAY | 12 | 195 | 6.797e-12 | 2.551e-10 |
| 125 | NEGATIVE REGULATION OF RESPONSE TO STIMULUS | 25 | 1360 | 7.184e-12 | 2.674e-10 |
| 126 | PHOSPHATIDYLINOSITOL 3 PHOSPHATE BIOSYNTHETIC PROCESS | 8 | 49 | 9.503e-12 | 3.509e-10 |
| 127 | REGULATION OF CATABOLIC PROCESS | 19 | 731 | 1.274e-11 | 4.669e-10 |
| 128 | NEGATIVE REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY IN ABSENCE OF LIGAND | 7 | 32 | 2.219e-11 | 8.002e-10 |
| 129 | NEGATIVE REGULATION OF SIGNAL TRANSDUCTION IN ABSENCE OF LIGAND | 7 | 32 | 2.219e-11 | 8.002e-10 |
| 130 | CELLULAR RESPONSE TO PEPTIDE | 13 | 274 | 2.24e-11 | 8.018e-10 |
| 131 | POSITIVE REGULATION OF NUCLEOCYTOPLASMIC TRANSPORT | 10 | 121 | 2.335e-11 | 8.294e-10 |
| 132 | NEGATIVE REGULATION OF CELL COMMUNICATION | 23 | 1192 | 2.419e-11 | 8.527e-10 |
| 133 | EXECUTION PHASE OF APOPTOSIS | 8 | 55 | 2.521e-11 | 8.819e-10 |
| 134 | DEFENSE RESPONSE | 23 | 1231 | 4.618e-11 | 1.603e-09 |
| 135 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 21 | 1004 | 5.044e-11 | 1.738e-09 |
| 136 | POSITIVE REGULATION OF BIOSYNTHETIC PROCESS | 27 | 1805 | 9.151e-11 | 3.131e-09 |
| 137 | NECROPTOTIC PROCESS | 6 | 21 | 1.083e-10 | 3.652e-09 |
| 138 | RESPONSE TO OXYGEN LEVELS | 13 | 311 | 1.083e-10 | 3.652e-09 |
| 139 | POSITIVE REGULATION OF NEURON DEATH | 8 | 67 | 1.304e-10 | 4.332e-09 |
| 140 | CELLULAR RESPONSE TO ORGANIC CYCLIC COMPOUND | 15 | 465 | 1.302e-10 | 4.332e-09 |
| 141 | CELLULAR RESPONSE TO HORMONE STIMULUS | 16 | 552 | 1.401e-10 | 4.624e-09 |
| 142 | STRESS ACTIVATED PROTEIN KINASE SIGNALING CASCADE | 9 | 103 | 1.495e-10 | 4.899e-09 |
| 143 | WOUND HEALING | 15 | 470 | 1.511e-10 | 4.915e-09 |
| 144 | RESPONSE TO WOUNDING | 16 | 563 | 1.871e-10 | 6.046e-09 |
| 145 | NEGATIVE REGULATION OF MOLECULAR FUNCTION | 21 | 1079 | 1.905e-10 | 6.114e-09 |
| 146 | CHEMICAL HOMEOSTASIS | 19 | 874 | 2.683e-10 | 8.55e-09 |
| 147 | REGULATION OF NECROPTOTIC PROCESS | 5 | 11 | 2.744e-10 | 8.687e-09 |
| 148 | RESPONSE TO AMINO ACID | 9 | 112 | 3.188e-10 | 1.002e-08 |
| 149 | CELLULAR GLUCOSE HOMEOSTASIS | 8 | 75 | 3.293e-10 | 1.028e-08 |
| 150 | PHOSPHATIDYLINOSITOL 3 KINASE SIGNALING | 6 | 25 | 3.494e-10 | 1.084e-08 |
| 151 | RESPONSE TO INTERLEUKIN 1 | 9 | 115 | 4.044e-10 | 1.246e-08 |
| 152 | CELLULAR RESPONSE TO BIOTIC STIMULUS | 10 | 163 | 4.494e-10 | 1.376e-08 |
| 153 | RESPONSE TO OXIDATIVE STRESS | 13 | 352 | 4.962e-10 | 1.509e-08 |
| 154 | REGULATION OF NUCLEOCYTOPLASMIC TRANSPORT | 11 | 220 | 5.059e-10 | 1.528e-08 |
| 155 | PHOSPHATIDYLINOSITOL BIOSYNTHETIC PROCESS | 9 | 120 | 5.926e-10 | 1.779e-08 |
| 156 | POSITIVE REGULATION OF PROTEIN SERINE THREONINE KINASE ACTIVITY | 12 | 289 | 6.521e-10 | 1.945e-08 |
| 157 | GLUCOSE HOMEOSTASIS | 10 | 170 | 6.784e-10 | 1.998e-08 |
| 158 | CARBOHYDRATE HOMEOSTASIS | 10 | 170 | 6.784e-10 | 1.998e-08 |
| 159 | POSITIVE REGULATION OF TRANSCRIPTION FACTOR IMPORT INTO NUCLEUS | 7 | 51 | 7.217e-10 | 2.112e-08 |
| 160 | NECROTIC CELL DEATH | 6 | 28 | 7.369e-10 | 2.143e-08 |
| 161 | NIK NF KAPPAB SIGNALING | 8 | 83 | 7.521e-10 | 2.174e-08 |
| 162 | INTERLEUKIN 1 MEDIATED SIGNALING PATHWAY | 5 | 13 | 7.601e-10 | 2.183e-08 |
| 163 | REGULATION OF INTRACELLULAR TRANSPORT | 16 | 621 | 7.796e-10 | 2.225e-08 |
| 164 | INOSITOL LIPID MEDIATED SIGNALING | 9 | 124 | 7.947e-10 | 2.255e-08 |
| 165 | POSITIVE REGULATION OF TRANSPORT | 19 | 936 | 8.447e-10 | 2.382e-08 |
| 166 | REGULATION OF PROTEIN INSERTION INTO MITOCHONDRIAL MEMBRANE INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 6 | 29 | 9.264e-10 | 2.581e-08 |
| 167 | POSITIVE REGULATION OF PROTEIN INSERTION INTO MITOCHONDRIAL MEMBRANE INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 6 | 29 | 9.264e-10 | 2.581e-08 |
| 168 | REGULATION OF PROTEIN LOCALIZATION | 19 | 950 | 1.081e-09 | 2.994e-08 |
| 169 | POSITIVE REGULATION OF PROTEIN LOCALIZATION TO NUCLEUS | 9 | 129 | 1.131e-09 | 3.114e-08 |
| 170 | CELLULAR RESPONSE TO INTERLEUKIN 1 | 8 | 88 | 1.208e-09 | 3.306e-08 |
| 171 | LIPOPOLYSACCHARIDE MEDIATED SIGNALING PATHWAY | 6 | 31 | 1.428e-09 | 3.884e-08 |
| 172 | T CELL APOPTOTIC PROCESS | 5 | 15 | 1.763e-09 | 4.771e-08 |
| 173 | RESPONSE TO ACID CHEMICAL | 12 | 319 | 2e-09 | 5.347e-08 |
| 174 | REGULATION OF MAP KINASE ACTIVITY | 12 | 319 | 2e-09 | 5.347e-08 |
| 175 | RESPONSE TO INORGANIC SUBSTANCE | 14 | 479 | 2.074e-09 | 5.515e-08 |
| 176 | REGULATION OF NEURON APOPTOTIC PROCESS | 10 | 192 | 2.217e-09 | 5.861e-08 |
| 177 | REGULATION OF TRANSCRIPTION FACTOR IMPORT INTO NUCLEUS | 8 | 95 | 2.237e-09 | 5.881e-08 |
| 178 | T CELL HOMEOSTASIS | 6 | 34 | 2.585e-09 | 6.757e-08 |
| 179 | POSITIVE REGULATION OF INTRACELLULAR PROTEIN TRANSPORT | 11 | 258 | 2.714e-09 | 7.056e-08 |
| 180 | ACTIVATION OF PROTEIN KINASE A ACTIVITY | 5 | 17 | 3.613e-09 | 9.339e-08 |
| 181 | POSITIVE REGULATION OF MITOCHONDRIAL OUTER MEMBRANE PERMEABILIZATION INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 6 | 36 | 3.722e-09 | 9.568e-08 |
| 182 | NEGATIVE REGULATION OF CATABOLIC PROCESS | 10 | 203 | 3.797e-09 | 9.706e-08 |
| 183 | PEPTIDYL SERINE MODIFICATION | 9 | 148 | 3.824e-09 | 9.723e-08 |
| 184 | POSITIVE REGULATION OF PROTEIN IMPORT | 8 | 104 | 4.617e-09 | 1.168e-07 |
| 185 | LYMPHOCYTE APOPTOTIC PROCESS | 5 | 18 | 4.988e-09 | 1.255e-07 |
| 186 | INNATE IMMUNE RESPONSE ACTIVATING CELL SURFACE RECEPTOR SIGNALING PATHWAY | 8 | 106 | 5.374e-09 | 1.344e-07 |
| 187 | REGULATION OF ORGANELLE ORGANIZATION | 20 | 1178 | 5.999e-09 | 1.493e-07 |
| 188 | NEGATIVE REGULATION OF CATALYTIC ACTIVITY | 17 | 829 | 6.661e-09 | 1.649e-07 |
| 189 | REGULATION OF LIPID METABOLIC PROCESS | 11 | 282 | 6.86e-09 | 1.689e-07 |
| 190 | REGULATION OF MEMBRANE PERMEABILITY | 7 | 70 | 7.067e-09 | 1.731e-07 |
| 191 | REGULATION OF PROTEIN LOCALIZATION TO NUCLEUS | 10 | 218 | 7.53e-09 | 1.834e-07 |
| 192 | CELLULAR RESPONSE TO CARBOHYDRATE STIMULUS | 7 | 74 | 1.049e-08 | 2.541e-07 |
| 193 | REGULATION OF INFLAMMATORY RESPONSE | 11 | 294 | 1.056e-08 | 2.546e-07 |
| 194 | REGULATION OF MITOCHONDRIAL OUTER MEMBRANE PERMEABILIZATION INVOLVED IN APOPTOTIC SIGNALING PATHWAY | 6 | 43 | 1.141e-08 | 2.723e-07 |
| 195 | CELLULAR COMPONENT DISASSEMBLY INVOLVED IN EXECUTION PHASE OF APOPTOSIS | 6 | 43 | 1.141e-08 | 2.723e-07 |
| 196 | RESPONSE TO CARBOHYDRATE | 9 | 168 | 1.163e-08 | 2.76e-07 |
| 197 | REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 24 | 1784 | 1.196e-08 | 2.825e-07 |
| 198 | REGULATION OF RELEASE OF CYTOCHROME C FROM MITOCHONDRIA | 6 | 44 | 1.318e-08 | 3.097e-07 |
| 199 | REGULATION OF INTRACELLULAR PROTEIN TRANSPORT | 12 | 381 | 1.457e-08 | 3.408e-07 |
| 200 | REGULATION OF MAPK CASCADE | 15 | 660 | 1.535e-08 | 3.571e-07 |
| 201 | POSITIVE REGULATION OF MAPK CASCADE | 13 | 470 | 1.604e-08 | 3.714e-07 |
| 202 | REGULATION OF MULTICELLULAR ORGANISMAL DEVELOPMENT | 23 | 1672 | 1.766e-08 | 4.068e-07 |
| 203 | CELL ACTIVATION | 14 | 568 | 1.802e-08 | 4.131e-07 |
| 204 | HEMOSTASIS | 11 | 311 | 1.885e-08 | 4.3e-07 |
| 205 | LEUKOCYTE APOPTOTIC PROCESS | 5 | 23 | 1.931e-08 | 4.383e-07 |
| 206 | RESPONSE TO TOXIC SUBSTANCE | 10 | 241 | 1.959e-08 | 4.424e-07 |
| 207 | POSITIVE REGULATION OF NEURON APOPTOTIC PROCESS | 6 | 47 | 1.987e-08 | 4.466e-07 |
| 208 | REGULATION OF EXECUTION PHASE OF APOPTOSIS | 5 | 24 | 2.432e-08 | 5.441e-07 |
| 209 | REGULATION OF PROTEIN IMPORT | 9 | 183 | 2.446e-08 | 5.446e-07 |
| 210 | RESPONSE TO VIRUS | 10 | 247 | 2.472e-08 | 5.478e-07 |
| 211 | SIGNAL TRANSDUCTION BY PROTEIN PHOSPHORYLATION | 12 | 404 | 2.783e-08 | 6.136e-07 |
| 212 | LYMPHOCYTE HOMEOSTASIS | 6 | 50 | 2.915e-08 | 6.398e-07 |
| 213 | RESPONSE TO NICOTINE | 6 | 51 | 3.294e-08 | 7.195e-07 |
| 214 | LEUKOCYTE CELL CELL ADHESION | 10 | 255 | 3.341e-08 | 7.265e-07 |
| 215 | NEGATIVE REGULATION OF CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY | 7 | 88 | 3.55e-08 | 7.647e-07 |
| 216 | RESPONSE TO REACTIVE OXYGEN SPECIES | 9 | 191 | 3.542e-08 | 7.647e-07 |
| 217 | PHOSPHATIDYLINOSITOL METABOLIC PROCESS | 9 | 193 | 3.875e-08 | 8.309e-07 |
| 218 | RESPONSE TO ALKALOID | 8 | 137 | 4.066e-08 | 8.678e-07 |
| 219 | AGING | 10 | 264 | 4.632e-08 | 9.842e-07 |
| 220 | MITOCHONDRIAL MEMBRANE ORGANIZATION | 7 | 92 | 4.842e-08 | 1.024e-06 |
| 221 | POSITIVE REGULATION OF NFAT PROTEIN IMPORT INTO NUCLEUS | 4 | 11 | 5.587e-08 | 1.176e-06 |
| 222 | RESPONSE TO DRUG | 12 | 431 | 5.651e-08 | 1.184e-06 |
| 223 | NEGATIVE REGULATION OF PROTEIN METABOLIC PROCESS | 18 | 1087 | 5.97e-08 | 1.246e-06 |
| 224 | REGULATION OF CELL DIFFERENTIATION | 21 | 1492 | 6.017e-08 | 1.25e-06 |
| 225 | REGULATION OF CELL ADHESION | 14 | 629 | 6.414e-08 | 1.327e-06 |
| 226 | POSITIVE REGULATION OF MAP KINASE ACTIVITY | 9 | 207 | 7.075e-08 | 1.457e-06 |
| 227 | REGULATION OF TRANSPORT | 23 | 1804 | 7.234e-08 | 1.483e-06 |
| 228 | LIPID PHOSPHORYLATION | 7 | 99 | 8.06e-08 | 1.638e-06 |
| 229 | REGULATION OF CELLULAR RESPONSE TO INSULIN STIMULUS | 6 | 59 | 8.051e-08 | 1.638e-06 |
| 230 | REGULATION OF ENDOTHELIAL CELL DEVELOPMENT | 4 | 12 | 8.356e-08 | 1.669e-06 |
| 231 | GLYCEROLIPID BIOSYNTHETIC PROCESS | 9 | 211 | 8.335e-08 | 1.669e-06 |
| 232 | REGULATION OF ESTABLISHMENT OF ENDOTHELIAL BARRIER | 4 | 12 | 8.356e-08 | 1.669e-06 |
| 233 | POSITIVE REGULATION OF GLUCOSE IMPORT IN RESPONSE TO INSULIN STIMULUS | 4 | 12 | 8.356e-08 | 1.669e-06 |
| 234 | POSITIVE REGULATION OF CYTOPLASMIC TRANSPORT | 10 | 282 | 8.594e-08 | 1.709e-06 |
| 235 | REGULATION OF GLUCOSE TRANSPORT | 7 | 100 | 8.641e-08 | 1.711e-06 |
| 236 | REGULATION OF GLUCOSE IMPORT | 6 | 60 | 8.919e-08 | 1.751e-06 |
| 237 | LEUKOCYTE HOMEOSTASIS | 6 | 60 | 8.919e-08 | 1.751e-06 |
| 238 | REGULATION OF REACTIVE OXYGEN SPECIES METABOLIC PROCESS | 8 | 152 | 9.121e-08 | 1.783e-06 |
| 239 | RENAL SYSTEM PROCESS | 7 | 102 | 9.911e-08 | 1.929e-06 |
| 240 | CELLULAR RESPONSE TO LIPID | 12 | 457 | 1.068e-07 | 2.07e-06 |
| 241 | REGULATION OF MITOCHONDRION ORGANIZATION | 9 | 218 | 1.102e-07 | 2.127e-06 |
| 242 | MYD88 DEPENDENT TOLL LIKE RECEPTOR SIGNALING PATHWAY | 5 | 32 | 1.126e-07 | 2.165e-06 |
| 243 | LEUKOCYTE DIFFERENTIATION | 10 | 292 | 1.189e-07 | 2.277e-06 |
| 244 | HEPATOCYTE APOPTOTIC PROCESS | 4 | 13 | 1.204e-07 | 2.286e-06 |
| 245 | RESPONSE TO COBALT ION | 4 | 13 | 1.204e-07 | 2.286e-06 |
| 246 | REGULATION OF CELLULAR LOCALIZATION | 19 | 1277 | 1.277e-07 | 2.416e-06 |
| 247 | REGULATION OF CYTOKINE PRODUCTION | 13 | 563 | 1.319e-07 | 2.485e-06 |
| 248 | PROTEIN KINASE B SIGNALING | 5 | 34 | 1.547e-07 | 2.879e-06 |
| 249 | RENAL WATER HOMEOSTASIS | 5 | 34 | 1.547e-07 | 2.879e-06 |
| 250 | CELLULAR RESPONSE TO ALKALOID | 5 | 34 | 1.547e-07 | 2.879e-06 |
| 251 | TOLL LIKE RECEPTOR 9 SIGNALING PATHWAY | 4 | 14 | 1.68e-07 | 3.115e-06 |
| 252 | REGULATION OF CYTOPLASMIC TRANSPORT | 12 | 481 | 1.854e-07 | 3.424e-06 |
| 253 | REGULATION OF PROTEIN TARGETING | 10 | 307 | 1.892e-07 | 3.48e-06 |
| 254 | IMMUNE SYSTEM DEVELOPMENT | 13 | 582 | 1.932e-07 | 3.538e-06 |
| 255 | INTRACELLULAR RECEPTOR SIGNALING PATHWAY | 8 | 168 | 1.973e-07 | 3.601e-06 |
| 256 | REGULATION OF CELL ACTIVATION | 12 | 484 | 1.982e-07 | 3.603e-06 |
| 257 | PHOSPHOLIPID BIOSYNTHETIC PROCESS | 9 | 235 | 2.085e-07 | 3.775e-06 |
| 258 | POSITIVE REGULATION OF CELL PROLIFERATION | 15 | 814 | 2.374e-07 | 4.281e-06 |
| 259 | CELLULAR RESPONSE TO GLUCAGON STIMULUS | 5 | 38 | 2.759e-07 | 4.956e-06 |
| 260 | RESPONSE TO CORTICOSTEROID | 8 | 176 | 2.818e-07 | 5.044e-06 |
| 261 | REGULATION OF RESPONSE TO WOUNDING | 11 | 413 | 3.315e-07 | 5.888e-06 |
| 262 | RESPONSE TO RADIATION | 11 | 413 | 3.315e-07 | 5.888e-06 |
| 263 | CELLULAR RESPONSE TO DNA DAMAGE STIMULUS | 14 | 720 | 3.338e-07 | 5.905e-06 |
| 264 | REGULATION OF AUTOPHAGY | 9 | 249 | 3.398e-07 | 5.967e-06 |
| 265 | REGULATION OF EPITHELIAL CELL DIFFERENTIATION | 7 | 122 | 3.39e-07 | 5.967e-06 |
| 266 | RESPONSE TO KETONE | 8 | 182 | 3.64e-07 | 6.368e-06 |
| 267 | REGULATION OF NFAT PROTEIN IMPORT INTO NUCLEUS | 4 | 17 | 3.962e-07 | 6.879e-06 |
| 268 | REGULATION OF GLUCOSE IMPORT IN RESPONSE TO INSULIN STIMULUS | 4 | 17 | 3.962e-07 | 6.879e-06 |
| 269 | RESPONSE TO METAL ION | 10 | 333 | 4e-07 | 6.918e-06 |
| 270 | RESPONSE TO UV | 7 | 126 | 4.223e-07 | 7.278e-06 |
| 271 | REGULATION OF NIK NF KAPPAB SIGNALING | 5 | 42 | 4.622e-07 | 7.907e-06 |
| 272 | POSITIVE REGULATION OF GLUCOSE TRANSPORT | 5 | 42 | 4.622e-07 | 7.907e-06 |
| 273 | LEUKOCYTE MIGRATION | 9 | 259 | 4.73e-07 | 8.062e-06 |
| 274 | POSITIVE REGULATION OF MULTICELLULAR ORGANISMAL PROCESS | 19 | 1395 | 5e-07 | 8.491e-06 |
| 275 | INSULIN RECEPTOR SIGNALING PATHWAY | 6 | 80 | 5.05e-07 | 8.545e-06 |
| 276 | LYMPHOCYTE ACTIVATION | 10 | 342 | 5.105e-07 | 8.606e-06 |
| 277 | PROTEIN OLIGOMERIZATION | 11 | 434 | 5.416e-07 | 9.097e-06 |
| 278 | JNK CASCADE | 6 | 82 | 5.851e-07 | 9.793e-06 |
| 279 | PROTEIN COMPLEX BIOGENESIS | 17 | 1132 | 5.95e-07 | 9.888e-06 |
| 280 | PROTEIN COMPLEX ASSEMBLY | 17 | 1132 | 5.95e-07 | 9.888e-06 |
| 281 | MULTICELLULAR ORGANISMAL HOMEOSTASIS | 9 | 272 | 7.123e-07 | 1.179e-05 |
| 282 | ACTIVATION OF MAPK ACTIVITY | 7 | 137 | 7.448e-07 | 1.229e-05 |
| 283 | POSITIVE REGULATION OF CELLULAR COMPONENT ORGANIZATION | 17 | 1152 | 7.591e-07 | 1.248e-05 |
| 284 | POSITIVE REGULATION OF REACTIVE OXYGEN SPECIES METABOLIC PROCESS | 6 | 86 | 7.765e-07 | 1.272e-05 |
| 285 | RESPONSE TO ANTIBIOTIC | 5 | 47 | 8.215e-07 | 1.341e-05 |
| 286 | RESPONSE TO ALCOHOL | 10 | 362 | 8.561e-07 | 1.393e-05 |
| 287 | RESPONSE TO GLUCAGON | 5 | 48 | 9.144e-07 | 1.483e-05 |
| 288 | SINGLE ORGANISM CELL ADHESION | 11 | 459 | 9.387e-07 | 1.517e-05 |
| 289 | NEGATIVE REGULATION OF LIPID CATABOLIC PROCESS | 4 | 21 | 9.852e-07 | 1.586e-05 |
| 290 | CELLULAR RESPONSE TO OXYGEN LEVELS | 7 | 143 | 9.945e-07 | 1.596e-05 |
| 291 | POSITIVE REGULATION OF CYTOKINE PRODUCTION | 10 | 370 | 1.043e-06 | 1.668e-05 |
| 292 | CELLULAR HOMEOSTASIS | 13 | 676 | 1.05e-06 | 1.674e-05 |
| 293 | NEGATIVE REGULATION OF IMMUNE SYSTEM PROCESS | 10 | 372 | 1.095e-06 | 1.74e-05 |
| 294 | RESPONSE TO GAMMA RADIATION | 5 | 50 | 1.125e-06 | 1.774e-05 |
| 295 | CELLULAR CHEMICAL HOMEOSTASIS | 12 | 570 | 1.122e-06 | 1.774e-05 |
| 296 | RESPONSE TO ESTRADIOL | 7 | 146 | 1.144e-06 | 1.798e-05 |
| 297 | POSITIVE REGULATION OF ORGANELLE ORGANIZATION | 12 | 573 | 1.185e-06 | 1.857e-05 |
| 298 | RELEASE OF CYTOCHROME C FROM MITOCHONDRIA | 4 | 22 | 1.201e-06 | 1.869e-05 |
| 299 | NEGATIVE REGULATION OF CELLULAR COMPONENT ORGANIZATION | 13 | 684 | 1.198e-06 | 1.869e-05 |
| 300 | RESPONSE TO TEMPERATURE STIMULUS | 7 | 148 | 1.253e-06 | 1.943e-05 |
| 301 | REGULATION OF CELL CELL ADHESION | 10 | 380 | 1.327e-06 | 2.052e-05 |
| 302 | POSITIVE REGULATION OF INTRINSIC APOPTOTIC SIGNALING PATHWAY | 5 | 52 | 1.372e-06 | 2.107e-05 |
| 303 | REGULATION OF LIPID CATABOLIC PROCESS | 5 | 52 | 1.372e-06 | 2.107e-05 |
| 304 | RESPONSE TO ESTROGEN | 8 | 218 | 1.424e-06 | 2.18e-05 |
| 305 | POSITIVE REGULATION OF CELLULAR RESPONSE TO INSULIN STIMULUS | 4 | 23 | 1.45e-06 | 2.211e-05 |
| 306 | GLYCEROPHOSPHOLIPID METABOLIC PROCESS | 9 | 297 | 1.477e-06 | 2.246e-05 |
| 307 | POSITIVE REGULATION OF EXTRINSIC APOPTOTIC SIGNALING PATHWAY | 5 | 53 | 1.511e-06 | 2.29e-05 |
| 308 | IMMUNE EFFECTOR PROCESS | 11 | 486 | 1.638e-06 | 2.474e-05 |
| 309 | REGULATION OF CELL CYCLE | 15 | 949 | 1.643e-06 | 2.474e-05 |
| 310 | MITOCHONDRION ORGANIZATION | 12 | 594 | 1.724e-06 | 2.588e-05 |
| 311 | REGULATION OF HOMOTYPIC CELL CELL ADHESION | 9 | 307 | 1.94e-06 | 2.902e-05 |
| 312 | NEGATIVE REGULATION OF HYDROLASE ACTIVITY | 10 | 397 | 1.966e-06 | 2.932e-05 |
| 313 | RESPONSE TO STEROID HORMONE | 11 | 497 | 2.034e-06 | 3.023e-05 |
| 314 | EPITHELIAL CELL APOPTOTIC PROCESS | 4 | 25 | 2.059e-06 | 3.051e-05 |
| 315 | APOPTOTIC MITOCHONDRIAL CHANGES | 5 | 57 | 2.179e-06 | 3.219e-05 |
| 316 | REGULATION OF LEUKOCYTE DIFFERENTIATION | 8 | 232 | 2.266e-06 | 3.337e-05 |
| 317 | REGULATION OF HEMOPOIESIS | 9 | 314 | 2.334e-06 | 3.426e-05 |
| 318 | MULTICELLULAR ORGANISMAL WATER HOMEOSTASIS | 5 | 58 | 2.378e-06 | 3.479e-05 |
| 319 | REGULATION OF BODY FLUID LEVELS | 11 | 506 | 2.418e-06 | 3.527e-05 |
| 320 | POSITIVE REGULATION OF MITOCHONDRION ORGANIZATION | 7 | 167 | 2.806e-06 | 4.08e-05 |
| 321 | REGULATION OF ENDOTHELIAL CELL DIFFERENTIATION | 4 | 27 | 2.841e-06 | 4.105e-05 |
| 322 | DNA CATABOLIC PROCESS | 4 | 27 | 2.841e-06 | 4.105e-05 |
| 323 | LEUKOCYTE ACTIVATION | 10 | 414 | 2.858e-06 | 4.117e-05 |
| 324 | RESPONSE TO HYDROGEN PEROXIDE | 6 | 109 | 3.128e-06 | 4.491e-05 |
| 325 | CELL DEVELOPMENT | 18 | 1426 | 3.175e-06 | 4.546e-05 |
| 326 | POSITIVE REGULATION OF CELL CELL ADHESION | 8 | 243 | 3.196e-06 | 4.562e-05 |
| 327 | POSITIVE REGULATION OF RELEASE OF CYTOCHROME C FROM MITOCHONDRIA | 4 | 28 | 3.305e-06 | 4.703e-05 |
| 328 | POSITIVE REGULATION OF NIK NF KAPPAB SIGNALING | 4 | 29 | 3.823e-06 | 5.423e-05 |
| 329 | PROTEIN HETEROOLIGOMERIZATION | 6 | 113 | 3.857e-06 | 5.455e-05 |
| 330 | MITOCHONDRIAL TRANSPORT | 7 | 177 | 4.124e-06 | 5.815e-05 |
| 331 | NEGATIVE REGULATION OF INTRACELLULAR SIGNAL TRANSDUCTION | 10 | 437 | 4.613e-06 | 6.485e-05 |
| 332 | REGULATION OF MYELOID CELL DIFFERENTIATION | 7 | 183 | 5.138e-06 | 7.2e-05 |
| 333 | INTRINSIC APOPTOTIC SIGNALING PATHWAY IN RESPONSE TO ENDOPLASMIC RETICULUM STRESS | 4 | 32 | 5.74e-06 | 8.02e-05 |
| 334 | WATER HOMEOSTASIS | 5 | 70 | 6.068e-06 | 8.453e-05 |
| 335 | CELL PROLIFERATION | 12 | 672 | 6.117e-06 | 8.496e-05 |
| 336 | MYELOID CELL DIFFERENTIATION | 7 | 189 | 6.35e-06 | 8.794e-05 |
| 337 | REGULATION OF RESPONSE TO EXTERNAL STIMULUS | 14 | 926 | 6.427e-06 | 8.874e-05 |
| 338 | GLYCEROLIPID METABOLIC PROCESS | 9 | 356 | 6.472e-06 | 8.91e-05 |
| 339 | PROTEIN AUTOPHOSPHORYLATION | 7 | 192 | 7.041e-06 | 9.664e-05 |
| 340 | NEGATIVE REGULATION OF PROTEIN SERINE THREONINE KINASE ACTIVITY | 6 | 126 | 7.239e-06 | 9.907e-05 |
| 341 | PHOSPHOLIPID METABOLIC PROCESS | 9 | 364 | 7.739e-06 | 0.0001056 |
| 342 | NEGATIVE REGULATION OF NECROTIC CELL DEATH | 3 | 11 | 7.854e-06 | 0.0001069 |
| 343 | REGULATION OF ESTABLISHMENT OF PROTEIN LOCALIZATION TO MITOCHONDRION | 6 | 128 | 7.925e-06 | 0.0001075 |
| 344 | PROTEIN COMPLEX SUBUNIT ORGANIZATION | 18 | 1527 | 8.249e-06 | 0.0001116 |
| 345 | POSITIVE REGULATION OF LEUKOCYTE DIFFERENTIATION | 6 | 131 | 9.053e-06 | 0.0001221 |
| 346 | ANATOMICAL STRUCTURE FORMATION INVOLVED IN MORPHOGENESIS | 14 | 957 | 9.347e-06 | 0.0001257 |
| 347 | RESPONSE TO GROWTH FACTOR | 10 | 475 | 9.578e-06 | 0.0001284 |
| 348 | MACROMOLECULAR COMPLEX ASSEMBLY | 17 | 1398 | 1.038e-05 | 0.0001388 |
| 349 | REGULATION OF VITAMIN METABOLIC PROCESS | 3 | 12 | 1.044e-05 | 0.0001392 |
| 350 | RESPONSE TO INSULIN | 7 | 205 | 1.08e-05 | 0.0001435 |
| 351 | NEGATIVE REGULATION OF LIPID METABOLIC PROCESS | 5 | 80 | 1.171e-05 | 0.0001552 |
| 352 | REGULATION OF PHOSPHATIDYLINOSITOL 3 KINASE SIGNALING | 6 | 138 | 1.22e-05 | 0.0001612 |
| 353 | LYMPHOCYTE DIFFERENTIATION | 7 | 209 | 1.224e-05 | 0.0001614 |
| 354 | LIPID MODIFICATION | 7 | 210 | 1.263e-05 | 0.000166 |
| 355 | REGULATION OF PROTEIN MATURATION | 5 | 82 | 1.322e-05 | 0.0001732 |
| 356 | POSITIVE REGULATION OF MACROPHAGE DIFFERENTIATION | 3 | 13 | 1.354e-05 | 0.0001765 |
| 357 | REGULATION OF BICELLULAR TIGHT JUNCTION ASSEMBLY | 3 | 13 | 1.354e-05 | 0.0001765 |
| 358 | CELL CELL ADHESION | 11 | 608 | 1.377e-05 | 0.000179 |
| 359 | REGULATION OF PHOSPHATIDYLINOSITOL 3 KINASE ACTIVITY | 4 | 40 | 1.426e-05 | 0.0001849 |
| 360 | PLATELET ACTIVATION | 6 | 142 | 1.435e-05 | 0.0001855 |
| 361 | GLAND DEVELOPMENT | 9 | 395 | 1.486e-05 | 0.0001915 |
| 362 | POSITIVE REGULATION OF DEVELOPMENTAL PROCESS | 15 | 1142 | 1.546e-05 | 0.0001987 |
| 363 | RESPONSE TO IONIZING RADIATION | 6 | 145 | 1.617e-05 | 0.0002067 |
| 364 | REGULATION OF INTRINSIC APOPTOTIC SIGNALING PATHWAY | 6 | 145 | 1.617e-05 | 0.0002067 |
| 365 | INNATE IMMUNE RESPONSE | 11 | 619 | 1.627e-05 | 0.0002074 |
| 366 | CELLULAR RESPONSE TO INSULIN STIMULUS | 6 | 146 | 1.681e-05 | 0.0002137 |
| 367 | REGULATION OF CAMP DEPENDENT PROTEIN KINASE ACTIVITY | 3 | 14 | 1.719e-05 | 0.0002179 |
| 368 | POSITIVE REGULATION OF PEPTIDYL SERINE PHOSPHORYLATION | 5 | 88 | 1.865e-05 | 0.0002358 |
| 369 | POSITIVE REGULATION OF CELL ACTIVATION | 8 | 311 | 1.933e-05 | 0.0002437 |
| 370 | RESPONSE TO HEAT | 5 | 89 | 1.97e-05 | 0.0002478 |
| 371 | APOPTOTIC DNA FRAGMENTATION | 3 | 15 | 2.143e-05 | 0.0002687 |
| 372 | SINGLE ORGANISM CELLULAR LOCALIZATION | 13 | 898 | 2.276e-05 | 0.0002847 |
| 373 | POSITIVE REGULATION OF INTERLEUKIN 8 PRODUCTION | 4 | 45 | 2.293e-05 | 0.0002861 |
| 374 | NEGATIVE REGULATION OF CELLULAR CATABOLIC PROCESS | 6 | 156 | 2.446e-05 | 0.0003043 |
| 375 | RESPONSE TO ENDOPLASMIC RETICULUM STRESS | 7 | 233 | 2.468e-05 | 0.0003062 |
| 376 | REGULATION OF CYTOKINE BIOSYNTHETIC PROCESS | 5 | 94 | 2.569e-05 | 0.0003179 |
| 377 | REGULATION OF LIPID TRANSPORT | 5 | 95 | 2.704e-05 | 0.0003337 |
| 378 | NEGATIVE REGULATION OF PROTEOLYSIS | 8 | 329 | 2.889e-05 | 0.0003556 |
| 379 | POSITIVE REGULATION OF REACTIVE OXYGEN SPECIES BIOSYNTHETIC PROCESS | 4 | 48 | 2.97e-05 | 0.0003637 |
| 380 | REGULATION OF LIPID KINASE ACTIVITY | 4 | 48 | 2.97e-05 | 0.0003637 |
| 381 | INTERSPECIES INTERACTION BETWEEN ORGANISMS | 11 | 662 | 3.019e-05 | 0.0003677 |
| 382 | SYMBIOSIS ENCOMPASSING MUTUALISM THROUGH PARASITISM | 11 | 662 | 3.019e-05 | 0.0003677 |
| 383 | POSITIVE REGULATION OF HEMOPOIESIS | 6 | 163 | 3.132e-05 | 0.0003805 |
| 384 | ACTIVATION OF NF KAPPAB INDUCING KINASE ACTIVITY | 3 | 17 | 3.185e-05 | 0.000385 |
| 385 | NEGATIVE REGULATION OF ANOIKIS | 3 | 17 | 3.185e-05 | 0.000385 |
| 386 | NEGATIVE REGULATION OF PEPTIDASE ACTIVITY | 7 | 245 | 3.401e-05 | 0.00041 |
| 387 | POSITIVE REGULATION OF MYELOID LEUKOCYTE DIFFERENTIATION | 4 | 50 | 3.496e-05 | 0.0004203 |
| 388 | NEGATIVE REGULATION OF ORGANIC ACID TRANSPORT | 3 | 18 | 3.812e-05 | 0.0004548 |
| 389 | REGULATION OF CELL CYCLE PROCESS | 10 | 558 | 3.809e-05 | 0.0004548 |
| 390 | INOSITOL PHOSPHATE MEDIATED SIGNALING | 3 | 18 | 3.812e-05 | 0.0004548 |
| 391 | NEGATIVE REGULATION OF KINASE ACTIVITY | 7 | 250 | 3.868e-05 | 0.0004603 |
| 392 | CELLULAR PROCESS INVOLVED IN REPRODUCTION IN MULTICELLULAR ORGANISM | 7 | 252 | 4.069e-05 | 0.0004829 |
| 393 | NEGATIVE REGULATION OF NEURON DEATH | 6 | 171 | 4.097e-05 | 0.0004851 |
| 394 | ORGANOPHOSPHATE BIOSYNTHETIC PROCESS | 9 | 450 | 4.13e-05 | 0.0004878 |
| 395 | STRIATED MUSCLE CELL DIFFERENTIATION | 6 | 173 | 4.372e-05 | 0.0005151 |
| 396 | REGULATION OF NITRIC OXIDE BIOSYNTHETIC PROCESS | 4 | 53 | 4.408e-05 | 0.0005166 |
| 397 | CELLULAR RESPONSE TO AMINO ACID STIMULUS | 4 | 53 | 4.408e-05 | 0.0005166 |
| 398 | REGULATION OF CELLULAR RESPONSE TO STRESS | 11 | 691 | 4.459e-05 | 0.0005213 |
| 399 | RESPONSE TO MUSCLE STRETCH | 3 | 19 | 4.515e-05 | 0.0005226 |
| 400 | NEGATIVE REGULATION OF MEIOTIC CELL CYCLE | 3 | 19 | 4.515e-05 | 0.0005226 |
| 401 | POSITIVE REGULATION OF PROTEIN EXPORT FROM NUCLEUS | 3 | 19 | 4.515e-05 | 0.0005226 |
| 402 | DNA CATABOLIC PROCESS ENDONUCLEOLYTIC | 3 | 19 | 4.515e-05 | 0.0005226 |
| 403 | NEGATIVE REGULATION OF TRANSFERASE ACTIVITY | 8 | 351 | 4.57e-05 | 0.0005263 |
| 404 | POSITIVE REGULATION OF CELL DIFFERENTIATION | 12 | 823 | 4.561e-05 | 0.0005263 |
| 405 | CELLULAR RESPONSE TO ACID CHEMICAL | 6 | 175 | 4.662e-05 | 0.0005343 |
| 406 | HOMEOSTASIS OF NUMBER OF CELLS | 6 | 175 | 4.662e-05 | 0.0005343 |
| 407 | REGULATION OF MYELOID LEUKOCYTE DIFFERENTIATION | 5 | 108 | 5.011e-05 | 0.0005729 |
| 408 | REGULATION OF MACROPHAGE DIFFERENTIATION | 3 | 20 | 5.298e-05 | 0.0006042 |
| 409 | POSITIVE REGULATION OF CYTOKINE BIOSYNTHETIC PROCESS | 4 | 58 | 6.297e-05 | 0.0007164 |
| 410 | GAMETE GENERATION | 10 | 595 | 6.525e-05 | 0.0007405 |
| 411 | POSITIVE REGULATION OF PROTEIN OLIGOMERIZATION | 3 | 22 | 7.119e-05 | 0.0008059 |
| 412 | REGULATION OF MONOOXYGENASE ACTIVITY | 4 | 60 | 7.198e-05 | 0.0008109 |
| 413 | DEVELOPMENTAL PROCESS INVOLVED IN REPRODUCTION | 10 | 602 | 7.191e-05 | 0.0008109 |
| 414 | POSITIVE REGULATION OF CELL ADHESION | 8 | 376 | 7.402e-05 | 0.0008319 |
| 415 | REGULATION OF PEPTIDYL SERINE PHOSPHORYLATION | 5 | 118 | 7.643e-05 | 0.0008569 |
| 416 | REGULATION OF PHOSPHOLIPID METABOLIC PROCESS | 4 | 61 | 7.681e-05 | 0.0008571 |
| 417 | REGULATION OF INTERLEUKIN 8 PRODUCTION | 4 | 61 | 7.681e-05 | 0.0008571 |
| 418 | RESPONSE TO LIGHT STIMULUS | 7 | 280 | 7.903e-05 | 0.0008797 |
| 419 | GLUCOSE METABOLIC PROCESS | 5 | 119 | 7.955e-05 | 0.0008834 |
| 420 | REGULATION OF B CELL ACTIVATION | 5 | 121 | 8.61e-05 | 0.0009538 |
| 421 | TRANSMEMBRANE RECEPTOR PROTEIN TYROSINE KINASE SIGNALING PATHWAY | 9 | 498 | 8.998e-05 | 0.0009944 |
| 422 | REGULATION OF ANOIKIS | 3 | 24 | 9.307e-05 | 0.001024 |
| 423 | T CELL DIFFERENTIATION | 5 | 123 | 9.304e-05 | 0.001024 |
| 424 | REGULATION OF REACTIVE OXYGEN SPECIES BIOSYNTHETIC PROCESS | 4 | 65 | 9.855e-05 | 0.001081 |
| 425 | EMBRYO DEVELOPMENT | 12 | 894 | 0.0001006 | 0.001102 |
| 426 | ESTABLISHMENT OF LOCALIZATION IN CELL | 17 | 1676 | 0.0001041 | 0.001138 |
| 427 | POSITIVE REGULATION OF LIPID BIOSYNTHETIC PROCESS | 4 | 66 | 0.0001046 | 0.00114 |
| 428 | CELLULAR EXTRAVASATION | 3 | 25 | 0.0001055 | 0.001144 |
| 429 | APOPTOTIC NUCLEAR CHANGES | 3 | 25 | 0.0001055 | 0.001144 |
| 430 | MEMBRANE ORGANIZATION | 12 | 899 | 0.0001061 | 0.001148 |
| 431 | CELLULAR RESPONSE TO DRUG | 4 | 67 | 0.0001109 | 0.001198 |
| 432 | REGULATION OF LIPID BIOSYNTHETIC PROCESS | 5 | 128 | 0.0001123 | 0.001207 |
| 433 | POSITIVE REGULATION OF LIPID METABOLIC PROCESS | 5 | 128 | 0.0001123 | 0.001207 |
| 434 | REGULATION OF LEUKOCYTE PROLIFERATION | 6 | 206 | 0.0001149 | 0.001229 |
| 435 | MULTICELLULAR ORGANISM REPRODUCTION | 11 | 768 | 0.0001146 | 0.001229 |
| 436 | CELLULAR COMPONENT DISASSEMBLY | 9 | 515 | 0.000116 | 0.001238 |
| 437 | NUCLEAR IMPORT | 5 | 129 | 0.0001165 | 0.00124 |
| 438 | NEGATIVE REGULATION OF NF KAPPAB TRANSCRIPTION FACTOR ACTIVITY | 4 | 68 | 0.0001176 | 0.001249 |
| 439 | DEVELOPMENTAL PROGRAMMED CELL DEATH | 3 | 26 | 0.0001189 | 0.001258 |
| 440 | NEGATIVE REGULATION OF LIPID TRANSPORT | 3 | 26 | 0.0001189 | 0.001258 |
| 441 | CELLULAR LIPID METABOLIC PROCESS | 12 | 913 | 0.0001227 | 0.001295 |
| 442 | B CELL ACTIVATION | 5 | 132 | 0.0001298 | 0.001367 |
| 443 | POSITIVE REGULATION OF TYPE I INTERFERON PRODUCTION | 4 | 70 | 0.0001316 | 0.001382 |
| 444 | REGULATION OF FATTY ACID TRANSPORT | 3 | 27 | 0.0001334 | 0.001398 |
| 445 | MACROMOLECULE CATABOLIC PROCESS | 12 | 926 | 0.0001401 | 0.001465 |
| 446 | POSITIVE REGULATION OF ACUTE INFLAMMATORY RESPONSE | 3 | 28 | 0.000149 | 0.001551 |
| 447 | MODULATION BY SYMBIONT OF HOST CELLULAR PROCESS | 3 | 28 | 0.000149 | 0.001551 |
| 448 | CELLULAR RESPONSE TO RADIATION | 5 | 137 | 0.0001545 | 0.001605 |
| 449 | POSITIVE REGULATION OF LOCOMOTION | 8 | 420 | 0.0001589 | 0.001646 |
| 450 | VASCULAR ENDOTHELIAL GROWTH FACTOR RECEPTOR SIGNALING PATHWAY | 4 | 74 | 0.0001633 | 0.001686 |
| 451 | LIPID BIOSYNTHETIC PROCESS | 9 | 539 | 0.0001634 | 0.001686 |
| 452 | PROTEIN IMPORT INTO NUCLEUS TRANSLOCATION | 3 | 29 | 0.0001658 | 0.001699 |
| 453 | MUSCLE ADAPTATION | 3 | 29 | 0.0001658 | 0.001699 |
| 454 | POSITIVE REGULATION OF MONOOXYGENASE ACTIVITY | 3 | 29 | 0.0001658 | 0.001699 |
| 455 | NEGATIVE REGULATION OF PHOSPHORUS METABOLIC PROCESS | 9 | 541 | 0.000168 | 0.001714 |
| 456 | NEGATIVE REGULATION OF PHOSPHATE METABOLIC PROCESS | 9 | 541 | 0.000168 | 0.001714 |
| 457 | POSITIVE REGULATION OF CARBOHYDRATE METABOLIC PROCESS | 4 | 75 | 0.000172 | 0.001748 |
| 458 | GRANULOCYTE MIGRATION | 4 | 75 | 0.000172 | 0.001748 |
| 459 | NEGATIVE REGULATION OF B CELL ACTIVATION | 3 | 30 | 0.0001837 | 0.001858 |
| 460 | REGULATION OF ANION TRANSMEMBRANE TRANSPORT | 3 | 30 | 0.0001837 | 0.001858 |
| 461 | NEGATIVE REGULATION OF CELL CYCLE | 8 | 433 | 0.0001955 | 0.001973 |
| 462 | EMBRYO DEVELOPMENT ENDING IN BIRTH OR EGG HATCHING | 9 | 554 | 0.0002005 | 0.002015 |
| 463 | LYMPHOCYTE COSTIMULATION | 4 | 78 | 0.0002002 | 0.002015 |
| 464 | REGULATION OF CELL CYCLE G1 S PHASE TRANSITION | 5 | 147 | 0.0002147 | 0.002153 |
| 465 | ENZYME LINKED RECEPTOR PROTEIN SIGNALING PATHWAY | 10 | 689 | 0.0002167 | 0.002168 |
| 466 | RESPONSE TO EXTRACELLULAR STIMULUS | 8 | 441 | 0.0002214 | 0.002207 |
| 467 | RESPONSE TO TRANSITION METAL NANOPARTICLE | 5 | 148 | 0.0002216 | 0.002207 |
| 468 | POSITIVE REGULATION OF MYELOID CELL DIFFERENTIATION | 4 | 81 | 0.0002316 | 0.002303 |
| 469 | REGULATION OF HOMEOSTATIC PROCESS | 8 | 447 | 0.0002425 | 0.002406 |
| 470 | NEGATIVE REGULATION OF MULTICELLULAR ORGANISMAL PROCESS | 12 | 983 | 0.0002437 | 0.002413 |
| 471 | REGULATION OF PROTEIN EXPORT FROM NUCLEUS | 3 | 33 | 0.0002449 | 0.002415 |
| 472 | NEGATIVE REGULATION OF ANION TRANSPORT | 3 | 33 | 0.0002449 | 0.002415 |
| 473 | MUSCLE CELL DIFFERENTIATION | 6 | 237 | 0.0002458 | 0.002418 |
| 474 | REPRODUCTION | 14 | 1297 | 0.0002518 | 0.002471 |
| 475 | PEPTIDYL AMINO ACID MODIFICATION | 11 | 841 | 0.0002526 | 0.002474 |
| 476 | RESPONSE TO FATTY ACID | 4 | 83 | 0.0002544 | 0.002487 |
| 477 | REGULATION OF DNA METABOLIC PROCESS | 7 | 340 | 0.0002615 | 0.002551 |
| 478 | PROTEIN IMPORT | 5 | 155 | 0.0002745 | 0.002672 |
| 479 | PROTEIN LOCALIZATION TO NUCLEUS | 5 | 156 | 0.0002827 | 0.002747 |
| 480 | NEGATIVE REGULATION OF TRANSPORT | 8 | 458 | 0.0002856 | 0.002769 |
| 481 | NEGATIVE REGULATION OF PROTEIN PROCESSING | 3 | 35 | 0.0002923 | 0.00281 |
| 482 | REGULATION OF PROTEIN OLIGOMERIZATION | 3 | 35 | 0.0002923 | 0.00281 |
| 483 | HEXOSE METABOLIC PROCESS | 5 | 157 | 0.0002912 | 0.00281 |
| 484 | NEGATIVE REGULATION OF PROTEIN MATURATION | 3 | 35 | 0.0002923 | 0.00281 |
| 485 | REGULATION OF JNK CASCADE | 5 | 159 | 0.0003087 | 0.002962 |
| 486 | NEGATIVE REGULATION OF GENE EXPRESSION | 15 | 1493 | 0.0003187 | 0.003051 |
| 487 | REGULATION OF MITOTIC CELL CYCLE | 8 | 468 | 0.0003301 | 0.003154 |
| 488 | REGULATION OF GENERATION OF PRECURSOR METABOLITES AND ENERGY | 4 | 89 | 0.0003324 | 0.003163 |
| 489 | EPITHELIAL CELL PROLIFERATION | 4 | 89 | 0.0003324 | 0.003163 |
| 490 | SEXUAL REPRODUCTION | 10 | 730 | 0.0003435 | 0.003262 |
| 491 | MODULATION BY VIRUS OF HOST MORPHOLOGY OR PHYSIOLOGY | 3 | 37 | 0.0003451 | 0.003271 |
| 492 | REGULATION OF OXIDOREDUCTASE ACTIVITY | 4 | 90 | 0.0003469 | 0.003274 |
| 493 | CALCIUM MEDIATED SIGNALING | 4 | 90 | 0.0003469 | 0.003274 |
| 494 | REGULATION OF PEPTIDE TRANSPORT | 6 | 256 | 0.0003712 | 0.003496 |
| 495 | ESTABLISHMENT OF PROTEIN LOCALIZATION TO ORGANELLE | 7 | 361 | 0.0003755 | 0.003529 |
| 496 | BIOLOGICAL ADHESION | 12 | 1032 | 0.0003795 | 0.00356 |
| 497 | ORGANOPHOSPHATE METABOLIC PROCESS | 11 | 885 | 0.0003901 | 0.003652 |
| 498 | WNT SIGNALING PATHWAY CALCIUM MODULATING PATHWAY | 3 | 39 | 0.0004038 | 0.003758 |
| 499 | ERBB2 SIGNALING PATHWAY | 3 | 39 | 0.0004038 | 0.003758 |
| 500 | SPLEEN DEVELOPMENT | 3 | 39 | 0.0004038 | 0.003758 |
| 501 | NEGATIVE REGULATION OF PROTEIN COMPLEX DISASSEMBLY | 5 | 170 | 0.0004197 | 0.003898 |
| 502 | FC GAMMA RECEPTOR SIGNALING PATHWAY | 4 | 95 | 0.0004262 | 0.00395 |
| 503 | REGULATION OF MEIOTIC CELL CYCLE | 3 | 40 | 0.0004354 | 0.00402 |
| 504 | REGULATION OF ACTIVATED T CELL PROLIFERATION | 3 | 40 | 0.0004354 | 0.00402 |
| 505 | NEGATIVE REGULATION OF PROTEIN MODIFICATION PROCESS | 9 | 616 | 0.0004368 | 0.004025 |
| 506 | MYELOID LEUKOCYTE DIFFERENTIATION | 4 | 96 | 0.0004435 | 0.004062 |
| 507 | SIGNAL TRANSDUCTION IN RESPONSE TO DNA DAMAGE | 4 | 96 | 0.0004435 | 0.004062 |
| 508 | FEMALE GAMETE GENERATION | 4 | 96 | 0.0004435 | 0.004062 |
| 509 | REGULATION OF LIPID STORAGE | 3 | 41 | 0.0004685 | 0.004266 |
| 510 | MITOTIC SPINDLE ASSEMBLY | 3 | 41 | 0.0004685 | 0.004266 |
| 511 | MICROTUBULE CYTOSKELETON ORGANIZATION INVOLVED IN MITOSIS | 3 | 41 | 0.0004685 | 0.004266 |
| 512 | MYELOID LEUKOCYTE MIGRATION | 4 | 99 | 0.0004984 | 0.004529 |
| 513 | REGULATION OF CELLULAR COMPONENT BIOGENESIS | 10 | 767 | 0.0005065 | 0.004594 |
| 514 | REGULATION OF TUMOR NECROSIS FACTOR SUPERFAMILY CYTOKINE PRODUCTION | 4 | 101 | 0.0005376 | 0.004865 |
| 515 | MYELOID LEUKOCYTE MEDIATED IMMUNITY | 3 | 43 | 0.0005395 | 0.004865 |
| 516 | REGULATION OF POLYSACCHARIDE METABOLIC PROCESS | 3 | 43 | 0.0005395 | 0.004865 |
| 517 | NEGATIVE REGULATION OF ORGANELLE ORGANIZATION | 7 | 387 | 0.0005684 | 0.005116 |
| 518 | REGULATION OF INTERLEUKIN 6 PRODUCTION | 4 | 104 | 0.0006004 | 0.005393 |
| 519 | CELLULAR RESPONSE TO OXIDATIVE STRESS | 5 | 184 | 0.0006017 | 0.005395 |
| 520 | THYMOCYTE AGGREGATION | 3 | 45 | 0.0006171 | 0.00549 |
| 521 | MODIFICATION BY SYMBIONT OF HOST MORPHOLOGY OR PHYSIOLOGY | 3 | 45 | 0.0006171 | 0.00549 |
| 522 | T CELL DIFFERENTIATION IN THYMUS | 3 | 45 | 0.0006171 | 0.00549 |
| 523 | REGULATION OF ALCOHOL BIOSYNTHETIC PROCESS | 3 | 45 | 0.0006171 | 0.00549 |
| 524 | MUSCLE SYSTEM PROCESS | 6 | 282 | 0.0006184 | 0.005491 |
| 525 | REGULATION OF PROTEIN CATABOLIC PROCESS | 7 | 393 | 0.0006225 | 0.005517 |
| 526 | PEPTIDYL THREONINE MODIFICATION | 3 | 46 | 0.0006584 | 0.005824 |
| 527 | REGULATION OF CELL CYCLE ARREST | 4 | 108 | 0.000692 | 0.00611 |
| 528 | SYSTEM PROCESS | 16 | 1785 | 0.0006939 | 0.006115 |
| 529 | PROTEIN DEPHOSPHORYLATION | 5 | 190 | 0.0006957 | 0.006119 |
| 530 | POSITIVE REGULATION OF OXIDOREDUCTASE ACTIVITY | 3 | 47 | 0.0007014 | 0.006158 |
| 531 | NEGATIVE REGULATION OF DEVELOPMENTAL PROCESS | 10 | 801 | 0.0007088 | 0.006211 |
| 532 | REGULATION OF CERAMIDE BIOSYNTHETIC PROCESS | 2 | 11 | 0.0007269 | 0.006334 |
| 533 | POSITIVE REGULATION OF HAIR CYCLE | 2 | 11 | 0.0007269 | 0.006334 |
| 534 | REGULATION OF FEVER GENERATION | 2 | 11 | 0.0007269 | 0.006334 |
| 535 | REGULATION OF NF KAPPAB IMPORT INTO NUCLEUS | 3 | 48 | 0.0007462 | 0.006478 |
| 536 | REGULATION OF ESTABLISHMENT OF PROTEIN LOCALIZATION TO PLASMA MEMBRANE | 3 | 48 | 0.0007462 | 0.006478 |
| 537 | REGULATION OF TYPE I INTERFERON PRODUCTION | 4 | 111 | 0.0007669 | 0.006645 |
| 538 | REGULATION OF TOLL LIKE RECEPTOR SIGNALING PATHWAY | 3 | 49 | 0.0007928 | 0.006844 |
| 539 | REGULATION OF STEROID BIOSYNTHETIC PROCESS | 3 | 49 | 0.0007928 | 0.006844 |
| 540 | REGULATION OF STRESS ACTIVATED PROTEIN KINASE SIGNALING CASCADE | 5 | 197 | 0.0008187 | 0.007052 |
| 541 | POSITIVE REGULATION OF INFLAMMATORY RESPONSE | 4 | 113 | 0.0008199 | 0.007052 |
| 542 | REGULATION OF ORGANIC ACID TRANSPORT | 3 | 50 | 0.0008411 | 0.007221 |
| 543 | REGULATION OF LYMPHOCYTE MEDIATED IMMUNITY | 4 | 114 | 0.0008474 | 0.007261 |
| 544 | REGULATION OF CHEMOKINE BIOSYNTHETIC PROCESS | 2 | 12 | 0.0008702 | 0.007389 |
| 545 | I KAPPAB PHOSPHORYLATION | 2 | 12 | 0.0008702 | 0.007389 |
| 546 | REPLICATIVE SENESCENCE | 2 | 12 | 0.0008702 | 0.007389 |
| 547 | MYELIN MAINTENANCE | 2 | 12 | 0.0008702 | 0.007389 |
| 548 | POSITIVE REGULATION OF INTERLEUKIN 2 BIOSYNTHETIC PROCESS | 2 | 12 | 0.0008702 | 0.007389 |
| 549 | NEGATIVE REGULATION OF I KAPPAB KINASE NF KAPPAB SIGNALING | 3 | 51 | 0.0008913 | 0.007541 |
| 550 | REGULATION OF INTERLEUKIN 12 PRODUCTION | 3 | 51 | 0.0008913 | 0.007541 |
| 551 | MONOSACCHARIDE METABOLIC PROCESS | 5 | 202 | 0.000916 | 0.007735 |
| 552 | PLASMA MEMBRANE ORGANIZATION | 5 | 203 | 0.0009365 | 0.007894 |
| 553 | CELLULAR RESPONSE TO IONIZING RADIATION | 3 | 52 | 0.0009434 | 0.007921 |
| 554 | REGULATION OF T CELL MEDIATED IMMUNITY | 3 | 52 | 0.0009434 | 0.007921 |
| 555 | NEGATIVE REGULATION OF PHOSPHORYLATION | 7 | 422 | 0.0009449 | 0.007921 |
| 556 | CELL MOTILITY | 10 | 835 | 0.0009738 | 0.00812 |
| 557 | LOCALIZATION OF CELL | 10 | 835 | 0.0009738 | 0.00812 |
| 558 | REGULATION OF IMMUNE EFFECTOR PROCESS | 7 | 424 | 0.0009712 | 0.00812 |
| 559 | REGULATION OF CELL DEVELOPMENT | 10 | 836 | 0.0009827 | 0.00818 |
| 560 | NEGATIVE REGULATION OF AUTOPHAGY | 3 | 53 | 0.0009974 | 0.008287 |
| 561 | REGULATION OF SPHINGOLIPID BIOSYNTHETIC PROCESS | 2 | 13 | 0.001026 | 0.00842 |
| 562 | REGULATION OF MEMBRANE LIPID METABOLIC PROCESS | 2 | 13 | 0.001026 | 0.00842 |
| 563 | CYTOKINE PRODUCTION | 4 | 120 | 0.001026 | 0.00842 |
| 564 | POSITIVE REGULATION OF STEROID BIOSYNTHETIC PROCESS | 2 | 13 | 0.001026 | 0.00842 |
| 565 | REGULATION OF HISTONE PHOSPHORYLATION | 2 | 13 | 0.001026 | 0.00842 |
| 566 | REGULATION OF PROTEIN POLYUBIQUITINATION | 2 | 13 | 0.001026 | 0.00842 |
| 567 | PROTEIN AUTOPROCESSING | 2 | 13 | 0.001026 | 0.00842 |
| 568 | NEGATIVE REGULATION OF REPRODUCTIVE PROCESS | 3 | 54 | 0.001053 | 0.008583 |
| 569 | REGULATION OF LYMPHOCYTE APOPTOTIC PROCESS | 3 | 54 | 0.001053 | 0.008583 |
| 570 | LIPID METABOLIC PROCESS | 12 | 1158 | 0.001051 | 0.008583 |
| 571 | REGULATION OF MITOCHONDRIAL MEMBRANE POTENTIAL | 3 | 54 | 0.001053 | 0.008583 |
| 572 | NEGATIVE REGULATION OF ESTABLISHMENT OF PROTEIN LOCALIZATION | 5 | 209 | 0.001066 | 0.008645 |
| 573 | REGULATION OF PEPTIDE SECRETION | 5 | 209 | 0.001066 | 0.008645 |
| 574 | GERM CELL DEVELOPMENT | 5 | 209 | 0.001066 | 0.008645 |
| 575 | POSITIVE REGULATION OF PROTEIN SECRETION | 5 | 211 | 0.001113 | 0.008987 |
| 576 | REGULATION OF B CELL PROLIFERATION | 3 | 55 | 0.001111 | 0.008987 |
| 577 | REGULATION OF ADAPTIVE IMMUNE RESPONSE | 4 | 123 | 0.001124 | 0.009067 |
| 578 | REGULATION OF FIBROBLAST APOPTOTIC PROCESS | 2 | 14 | 0.001194 | 0.009563 |
| 579 | INSULIN LIKE GROWTH FACTOR RECEPTOR SIGNALING PATHWAY | 2 | 14 | 0.001194 | 0.009563 |
| 580 | T CELL MIGRATION | 2 | 14 | 0.001194 | 0.009563 |
| 581 | DETERMINATION OF ADULT LIFESPAN | 2 | 14 | 0.001194 | 0.009563 |
| 582 | TISSUE DEVELOPMENT | 14 | 1518 | 0.001206 | 0.009639 |
| 583 | POSITIVE REGULATION OF HOMEOSTATIC PROCESS | 5 | 216 | 0.001234 | 0.009835 |
| 584 | POSITIVE REGULATION OF TUMOR NECROSIS FACTOR SUPERFAMILY CYTOKINE PRODUCTION | 3 | 57 | 0.001233 | 0.009835 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | TUMOR NECROSIS FACTOR RECEPTOR SUPERFAMILY BINDING | 12 | 47 | 1.217e-19 | 1.13e-16 |
| 2 | DEATH RECEPTOR BINDING | 9 | 18 | 3.716e-18 | 1.726e-15 |
| 3 | CYTOKINE RECEPTOR BINDING | 17 | 271 | 1.216e-16 | 3.767e-14 |
| 4 | ENZYME BINDING | 33 | 1737 | 2.377e-16 | 5.521e-14 |
| 5 | KINASE ACTIVITY | 24 | 842 | 1.654e-15 | 3.072e-13 |
| 6 | TRANSFERASE ACTIVITY TRANSFERRING PHOSPHORUS CONTAINING GROUPS | 24 | 992 | 6.11e-14 | 9.46e-12 |
| 7 | PROTEIN SERINE THREONINE KINASE ACTIVITY | 17 | 445 | 4.249e-13 | 5.639e-11 |
| 8 | UBIQUITIN LIKE PROTEIN LIGASE BINDING | 14 | 264 | 7.775e-13 | 9.029e-11 |
| 9 | X1 PHOSPHATIDYLINOSITOL 3 KINASE ACTIVITY | 8 | 43 | 3.11e-12 | 3.21e-10 |
| 10 | PHOSPHATIDYLINOSITOL 3 KINASE ACTIVITY | 9 | 70 | 4.268e-12 | 3.965e-10 |
| 11 | CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY INVOLVED IN APOPTOTIC PROCESS | 6 | 15 | 1.017e-11 | 8.587e-10 |
| 12 | PROTEIN HETERODIMERIZATION ACTIVITY | 16 | 468 | 1.213e-11 | 8.858e-10 |
| 13 | PROTEIN KINASE ACTIVITY | 18 | 640 | 1.335e-11 | 8.858e-10 |
| 14 | PHOSPHATIDYLINOSITOL KINASE ACTIVITY | 8 | 51 | 1.334e-11 | 8.858e-10 |
| 15 | PROTEASE BINDING | 9 | 104 | 1.632e-10 | 1.011e-08 |
| 16 | ADENYL NUCLEOTIDE BINDING | 24 | 1514 | 4.557e-10 | 2.646e-08 |
| 17 | KINASE BINDING | 16 | 606 | 5.471e-10 | 2.99e-08 |
| 18 | CYSTEINE TYPE ENDOPEPTIDASE ACTIVITY | 8 | 86 | 1.003e-09 | 5.176e-08 |
| 19 | TUMOR NECROSIS FACTOR RECEPTOR BINDING | 6 | 30 | 1.155e-09 | 5.645e-08 |
| 20 | IDENTICAL PROTEIN BINDING | 20 | 1209 | 9.312e-09 | 3.952e-07 |
| 21 | ENZYME REGULATOR ACTIVITY | 18 | 959 | 8.752e-09 | 3.952e-07 |
| 22 | RECEPTOR BINDING | 22 | 1476 | 9.359e-09 | 3.952e-07 |
| 23 | CYSTEINE TYPE ENDOPEPTIDASE REGULATOR ACTIVITY INVOLVED IN APOPTOTIC PROCESS | 6 | 42 | 9.85e-09 | 3.978e-07 |
| 24 | PROTEIN DIMERIZATION ACTIVITY | 19 | 1149 | 2.404e-08 | 9.038e-07 |
| 25 | DEATH RECEPTOR ACTIVITY | 5 | 24 | 2.432e-08 | 9.038e-07 |
| 26 | RIBONUCLEOTIDE BINDING | 24 | 1860 | 2.688e-08 | 9.603e-07 |
| 27 | KINASE REGULATOR ACTIVITY | 9 | 186 | 2.816e-08 | 9.689e-07 |
| 28 | PROTEIN DOMAIN SPECIFIC BINDING | 14 | 624 | 5.812e-08 | 1.928e-06 |
| 29 | MOLECULAR FUNCTION REGULATOR | 20 | 1353 | 6.081e-08 | 1.948e-06 |
| 30 | PROTEIN PHOSPHATASE BINDING | 7 | 120 | 3.028e-07 | 8.82e-06 |
| 31 | INTERLEUKIN 1 RECEPTOR BINDING | 4 | 16 | 3.038e-07 | 8.82e-06 |
| 32 | PHOSPHATIDYLINOSITOL PHOSPHATE KINASE ACTIVITY | 4 | 16 | 3.038e-07 | 8.82e-06 |
| 33 | CYSTEINE TYPE PEPTIDASE ACTIVITY | 8 | 184 | 3.956e-07 | 1.114e-05 |
| 34 | PHOSPHATASE BINDING | 7 | 162 | 2.293e-06 | 6.264e-05 |
| 35 | PROTEIN SERINE THREONINE KINASE INHIBITOR ACTIVITY | 4 | 30 | 4.399e-06 | 0.0001168 |
| 36 | INSULIN RECEPTOR SUBSTRATE BINDING | 3 | 11 | 7.854e-06 | 0.0002027 |
| 37 | PEPTIDASE REGULATOR ACTIVITY | 7 | 214 | 1.427e-05 | 0.0003583 |
| 38 | PROTEIN KINASE A BINDING | 4 | 42 | 1.737e-05 | 0.0004247 |
| 39 | PROTEIN KINASE A CATALYTIC SUBUNIT BINDING | 3 | 15 | 2.143e-05 | 0.0005104 |
| 40 | PROTEIN COMPLEX BINDING | 13 | 935 | 3.466e-05 | 0.0008051 |
| 41 | PEPTIDASE ACTIVATOR ACTIVITY INVOLVED IN APOPTOTIC PROCESS | 3 | 21 | 6.164e-05 | 0.001397 |
| 42 | ENZYME INHIBITOR ACTIVITY | 8 | 378 | 7.68e-05 | 0.001699 |
| 43 | CAMP BINDING | 3 | 23 | 8.165e-05 | 0.001724 |
| 44 | CYSTEINE TYPE ENDOPEPTIDASE INHIBITOR ACTIVITY INVOLVED IN APOPTOTIC PROCESS | 3 | 23 | 8.165e-05 | 0.001724 |
| 45 | PROTEIN SERINE THREONINE PHOSPHATASE ACTIVITY | 4 | 64 | 9.274e-05 | 0.001915 |
| 46 | GROWTH FACTOR RECEPTOR BINDING | 5 | 129 | 0.0001165 | 0.002353 |
| 47 | PROTEIN PHOSPHATASE 2A BINDING | 3 | 28 | 0.000149 | 0.002946 |
| 48 | SIGNAL TRANSDUCER ACTIVITY | 17 | 1731 | 0.0001544 | 0.002979 |
| 49 | MACROMOLECULAR COMPLEX BINDING | 15 | 1399 | 0.0001571 | 0.002979 |
| 50 | ENDOPEPTIDASE ACTIVITY | 8 | 448 | 0.0002462 | 0.004575 |
| 51 | PROTEIN HOMODIMERIZATION ACTIVITY | 10 | 722 | 0.0003148 | 0.005681 |
| 52 | CYCLIC NUCLEOTIDE BINDING | 3 | 36 | 0.000318 | 0.005681 |
| 53 | HEAT SHOCK PROTEIN BINDING | 4 | 89 | 0.0003324 | 0.005718 |
| 54 | KINASE INHIBITOR ACTIVITY | 4 | 89 | 0.0003324 | 0.005718 |
| 55 | PEPTIDASE ACTIVATOR ACTIVITY | 3 | 38 | 0.0003737 | 0.006313 |
| 56 | PROTEIN N TERMINUS BINDING | 4 | 103 | 0.0005789 | 0.009603 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | TRANSFERASE COMPLEX TRANSFERRING PHOSPHORUS CONTAINING GROUPS | 17 | 237 | 1.267e-17 | 4.282e-15 |
| 2 | MEMBRANE MICRODOMAIN | 18 | 288 | 1.466e-17 | 4.282e-15 |
| 3 | CATALYTIC COMPLEX | 26 | 1038 | 1.747e-15 | 3.401e-13 |
| 4 | PHOSPHATIDYLINOSITOL 3 KINASE COMPLEX | 8 | 20 | 2.891e-15 | 4.22e-13 |
| 5 | MEMBRANE PROTEIN COMPLEX | 23 | 1020 | 1.008e-12 | 1.177e-10 |
| 6 | TRANSFERASE COMPLEX | 17 | 703 | 5.563e-10 | 5.415e-08 |
| 7 | PROTEIN KINASE COMPLEX | 8 | 90 | 1.448e-09 | 1.208e-07 |
| 8 | CYTOSOLIC PART | 10 | 223 | 9.353e-09 | 6.828e-07 |
| 9 | CILIARY BASE | 5 | 23 | 1.931e-08 | 1.137e-06 |
| 10 | MEMBRANE REGION | 19 | 1134 | 1.948e-08 | 1.137e-06 |
| 11 | PLASMA MEMBRANE PROTEIN COMPLEX | 13 | 510 | 4.19e-08 | 2.224e-06 |
| 12 | EXTRINSIC COMPONENT OF MEMBRANE | 8 | 252 | 4.181e-06 | 0.0002035 |
| 13 | MITOCHONDRION | 19 | 1633 | 5.238e-06 | 0.0002353 |
| 14 | CD40 RECEPTOR COMPLEX | 3 | 11 | 7.854e-06 | 0.0003058 |
| 15 | IKAPPAB KINASE COMPLEX | 3 | 11 | 7.854e-06 | 0.0003058 |
| 16 | RECEPTOR COMPLEX | 8 | 327 | 2.766e-05 | 0.00101 |
| 17 | PHOSPHATASE COMPLEX | 4 | 48 | 2.97e-05 | 0.00102 |
| 18 | PLASMA MEMBRANE RAFT | 4 | 86 | 0.0002915 | 0.009458 |
Over-represented Pathway
| Num | Pathway | Pathview | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|---|
| 1 | hsa04210_Apoptosis | 74 | 89 | 7.651e-195 | 1.354e-192 | |
| 2 | hsa04620_Toll.like_receptor_signaling_pathway | 25 | 102 | 5.854e-40 | 5.181e-38 | |
| 3 | hsa04380_Osteoclast_differentiation | 25 | 128 | 3.237e-37 | 1.91e-35 | |
| 4 | hsa04722_Neurotrophin_signaling_pathway | 24 | 127 | 2.514e-35 | 1.112e-33 | |
| 5 | hsa04660_T_cell_receptor_signaling_pathway | 23 | 108 | 3.801e-35 | 1.346e-33 | |
| 6 | hsa04662_B_cell_receptor_signaling_pathway | 21 | 75 | 6.839e-35 | 2.017e-33 | |
| 7 | hsa04010_MAPK_signaling_pathway | 26 | 268 | 2.202e-30 | 5.567e-29 | |
| 8 | hsa04650_Natural_killer_cell_mediated_cytotoxicity | 21 | 136 | 6.816e-29 | 1.508e-27 | |
| 9 | hsa04370_VEGF_signaling_pathway | 17 | 76 | 2.044e-26 | 4.019e-25 | |
| 10 | hsa04910_Insulin_signaling_pathway | 19 | 138 | 3.741e-25 | 6.622e-24 | |
| 11 | hsa04062_Chemokine_signaling_pathway | 20 | 189 | 4.371e-24 | 7.034e-23 | |
| 12 | hsa04914_Progesterone.mediated_oocyte_maturation | 14 | 87 | 1.074e-19 | 1.584e-18 | |
| 13 | hsa04510_Focal_adhesion | 17 | 200 | 6.994e-19 | 9.523e-18 | |
| 14 | hsa04621_NOD.like_receptor_signaling_pathway | 12 | 59 | 2.519e-18 | 3.184e-17 | |
| 15 | hsa04973_Carbohydrate_digestion_and_absorption | 11 | 44 | 5.694e-18 | 6.719e-17 | |
| 16 | hsa04920_Adipocytokine_signaling_pathway | 12 | 68 | 1.597e-17 | 1.766e-16 | |
| 17 | hsa04622_RIG.I.like_receptor_signaling_pathway | 12 | 71 | 2.783e-17 | 2.897e-16 | |
| 18 | hsa04150_mTOR_signaling_pathway | 11 | 52 | 4.382e-17 | 4.309e-16 | |
| 19 | hsa04664_Fc_epsilon_RI_signaling_pathway | 12 | 79 | 1.088e-16 | 1.013e-15 | |
| 20 | hsa04012_ErbB_signaling_pathway | 12 | 87 | 3.682e-16 | 3.259e-15 | |
| 21 | hsa04630_Jak.STAT_signaling_pathway | 13 | 155 | 1.522e-14 | 1.282e-13 | |
| 22 | hsa04666_Fc_gamma_R.mediated_phagocytosis | 11 | 95 | 5.001e-14 | 4.024e-13 | |
| 23 | hsa04115_p53_signaling_pathway | 10 | 69 | 7.305e-14 | 5.621e-13 | |
| 24 | hsa04960_Aldosterone.regulated_sodium_reabsorption | 8 | 42 | 2.539e-12 | 1.872e-11 | |
| 25 | hsa04623_Cytosolic_DNA.sensing_pathway | 8 | 56 | 2.932e-11 | 2.076e-10 | |
| 26 | hsa04070_Phosphatidylinositol_signaling_system | 8 | 78 | 4.537e-10 | 3.089e-09 | |
| 27 | hsa04720_Long.term_potentiation | 7 | 70 | 7.067e-09 | 4.633e-08 | |
| 28 | hsa04670_Leukocyte_transendothelial_migration | 8 | 117 | 1.177e-08 | 7.437e-08 | |
| 29 | hsa04310_Wnt_signaling_pathway | 8 | 151 | 8.666e-08 | 5.289e-07 | |
| 30 | hsa04114_Oocyte_meiosis | 7 | 114 | 2.132e-07 | 1.258e-06 | |
| 31 | hsa04810_Regulation_of_actin_cytoskeleton | 8 | 214 | 1.239e-06 | 7.077e-06 | |
| 32 | hsa04020_Calcium_signaling_pathway | 7 | 177 | 4.124e-06 | 2.281e-05 | |
| 33 | hsa00562_Inositol_phosphate_metabolism | 4 | 57 | 5.88e-05 | 0.0003154 | |
| 34 | hsa04640_Hematopoietic_cell_lineage | 4 | 88 | 0.0003183 | 0.001657 | |
| 35 | hsa04962_Vasopressin.regulated_water_reabsorption | 3 | 44 | 0.0005775 | 0.00292 | |
| 36 | hsa04742_Taste_transduction | 3 | 52 | 0.0009434 | 0.004638 | |
| 37 | hsa04340_Hedgehog_signaling_pathway | 3 | 56 | 0.001171 | 0.005601 | |
| 38 | hsa04360_Axon_guidance | 4 | 130 | 0.00138 | 0.006427 | |
| 39 | hsa04976_Bile_secretion | 3 | 71 | 0.00232 | 0.01053 | |
| 40 | hsa04971_Gastric_acid_secretion | 3 | 74 | 0.002611 | 0.01155 | |
| 41 | hsa04970_Salivary_secretion | 3 | 89 | 0.004397 | 0.01898 | |
| 42 | hsa04540_Gap_junction | 3 | 90 | 0.004536 | 0.01912 | |
| 43 | hsa04912_GnRH_signaling_pathway | 3 | 101 | 0.006251 | 0.02515 | |
| 44 | hsa04916_Melanogenesis | 3 | 101 | 0.006251 | 0.02515 | |
| 45 | hsa04270_Vascular_smooth_muscle_contraction | 3 | 116 | 0.00914 | 0.03595 | |
| 46 | hsa04530_Tight_junction | 3 | 133 | 0.01322 | 0.05087 | |
| 47 | hsa04120_Ubiquitin_mediated_proteolysis | 3 | 139 | 0.01487 | 0.05601 | |
| 48 | hsa04141_Protein_processing_in_endoplasmic_reticulum | 3 | 168 | 0.02444 | 0.09011 | |
| 49 | hsa04110_Cell_cycle | 2 | 128 | 0.08151 | 0.2944 | |
| 50 | hsa04740_Olfactory_transduction | 3 | 388 | 0.1735 | 0.6023 |
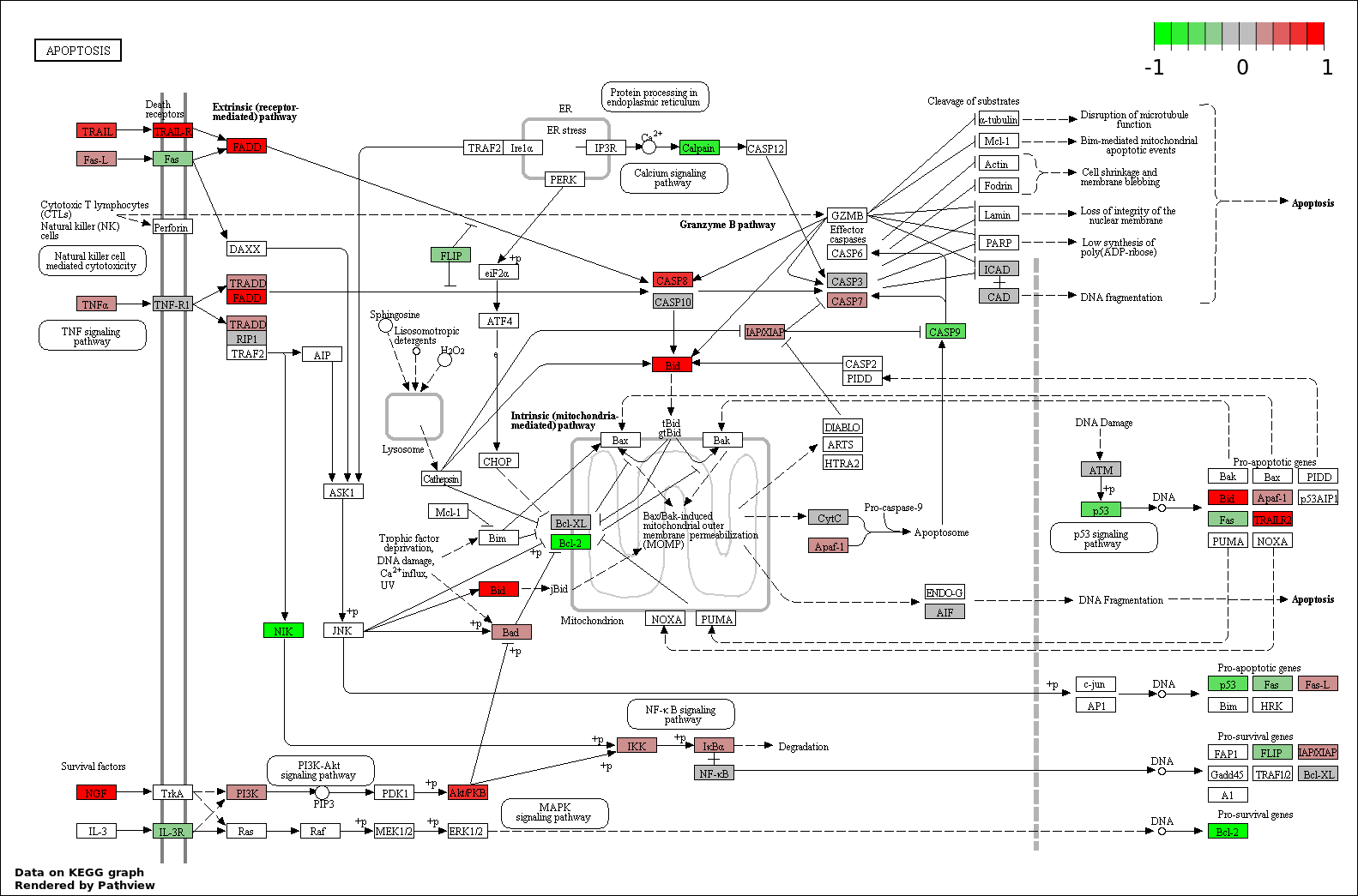
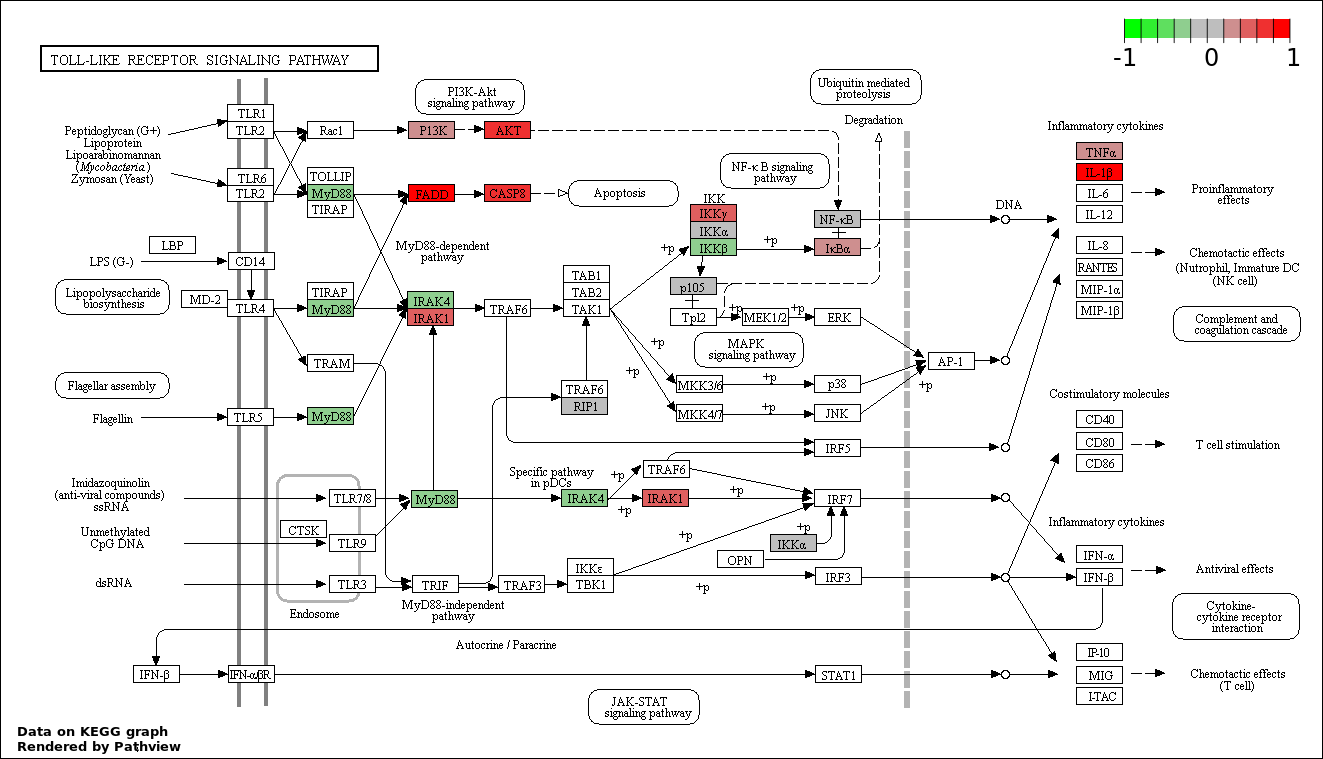
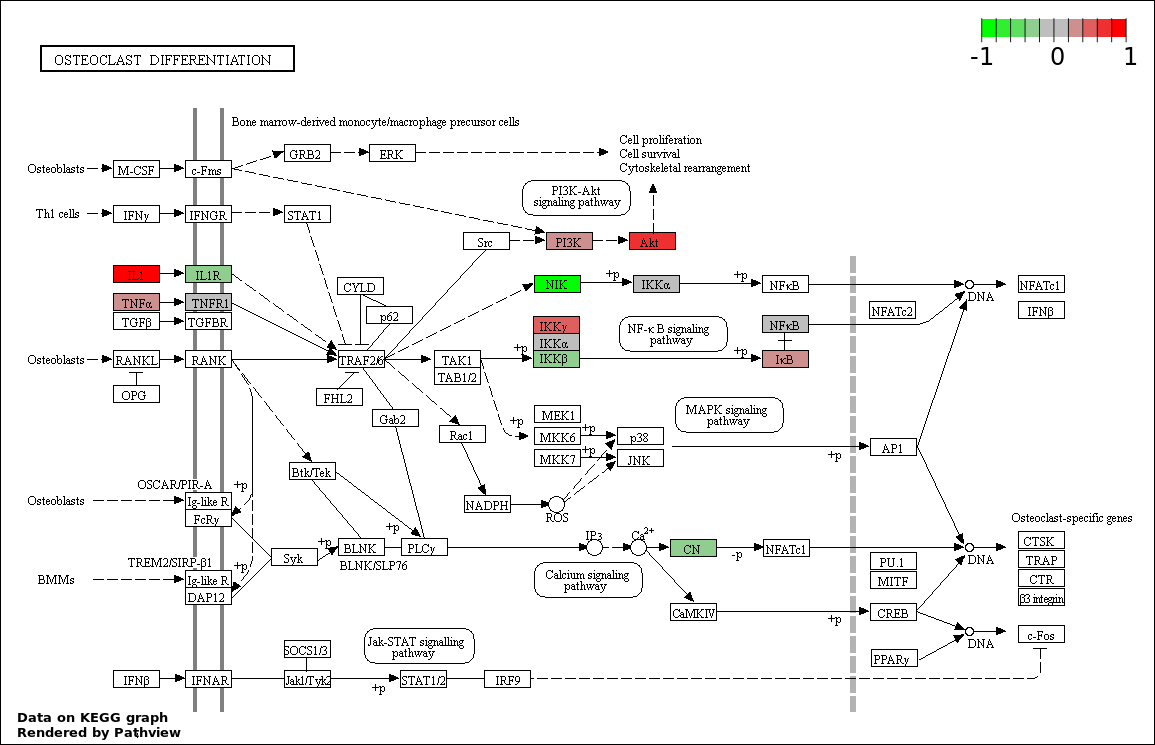
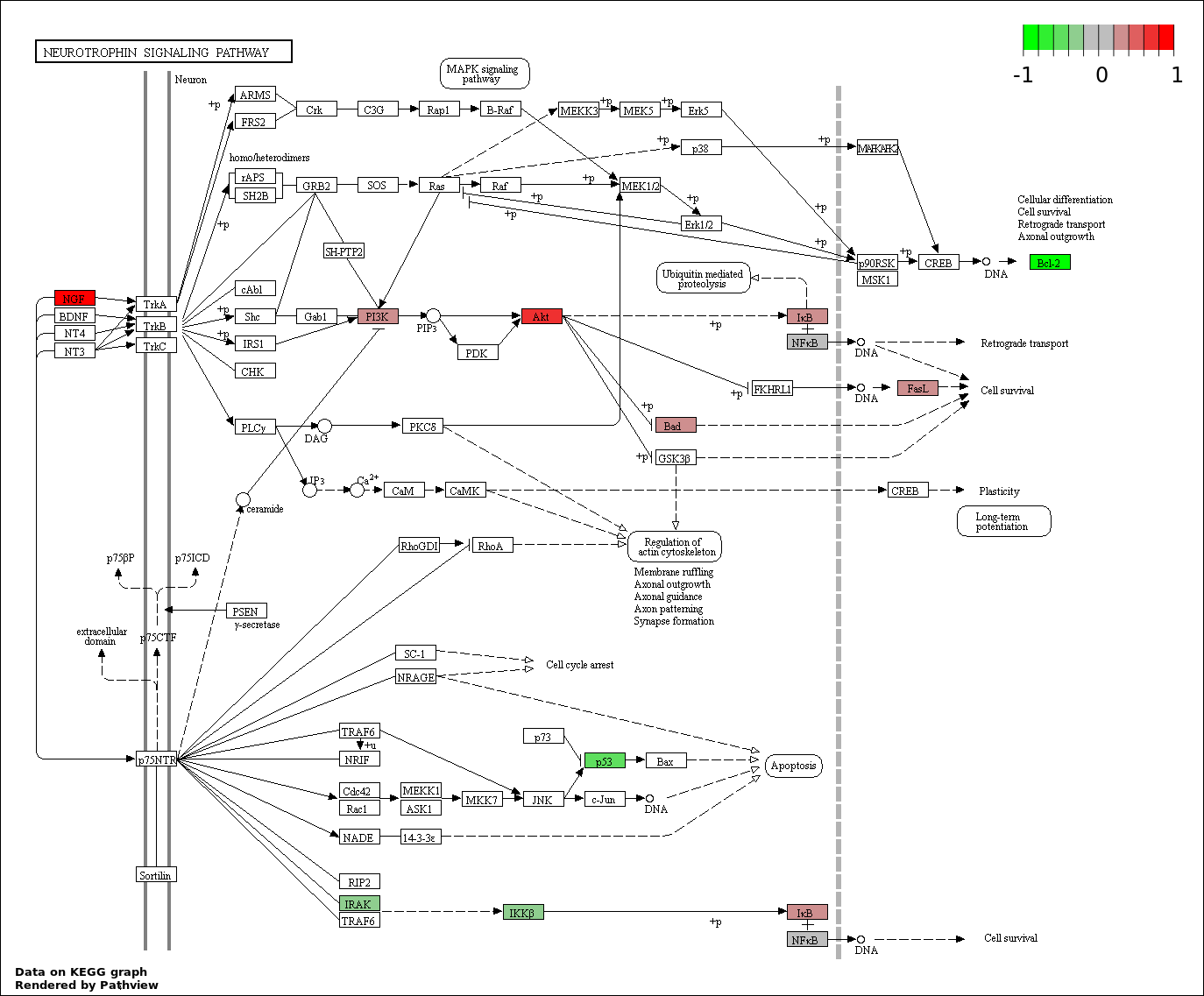
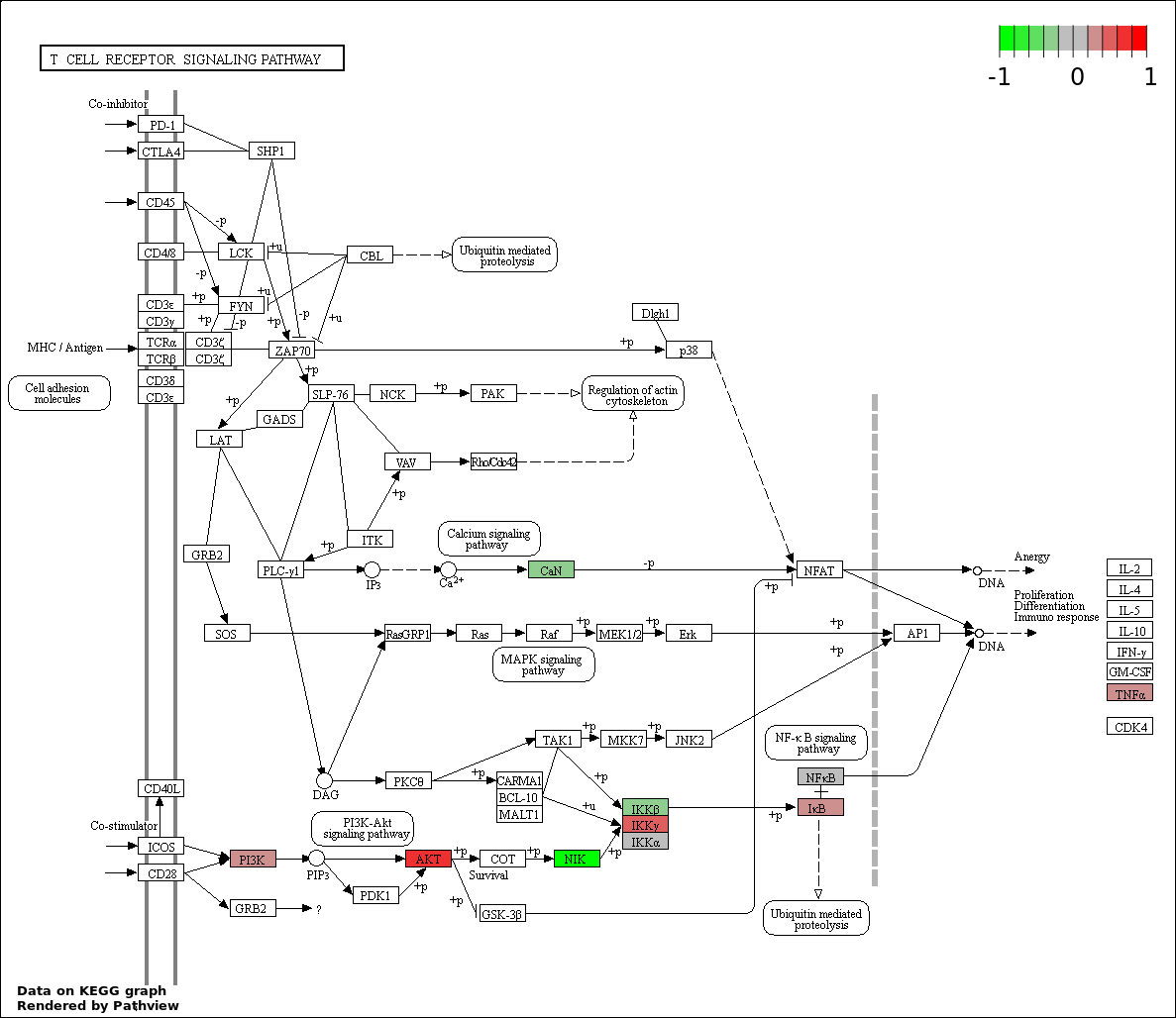
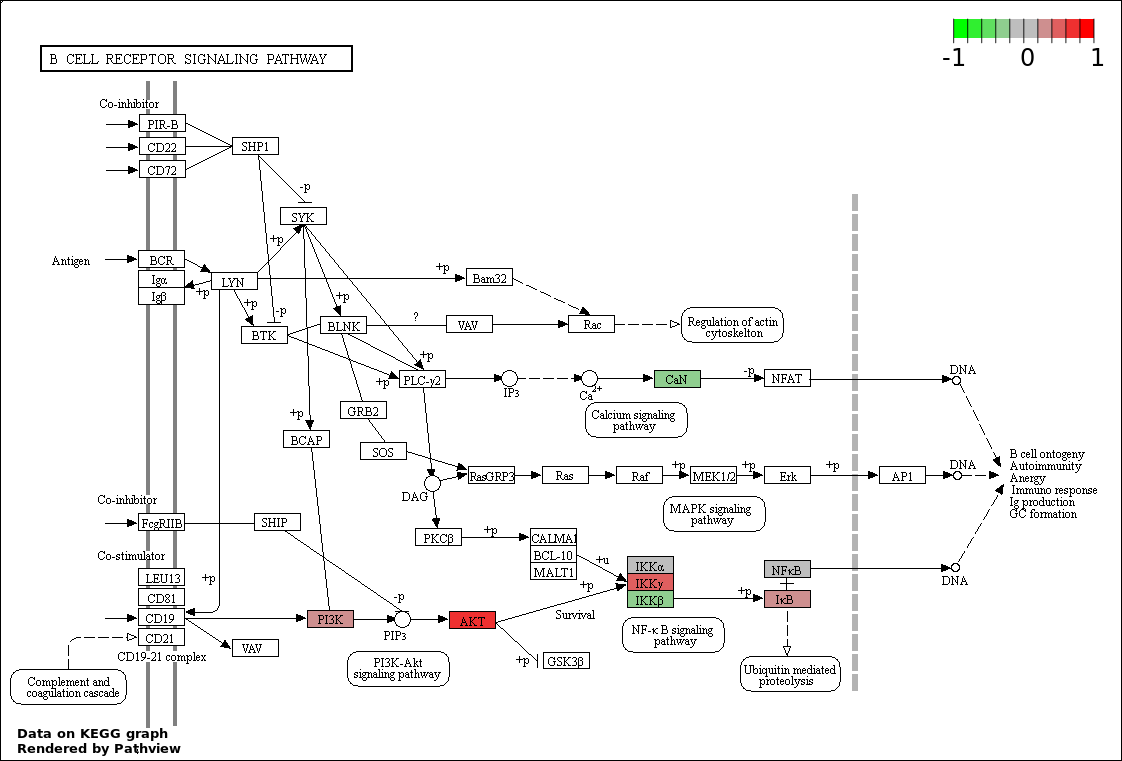
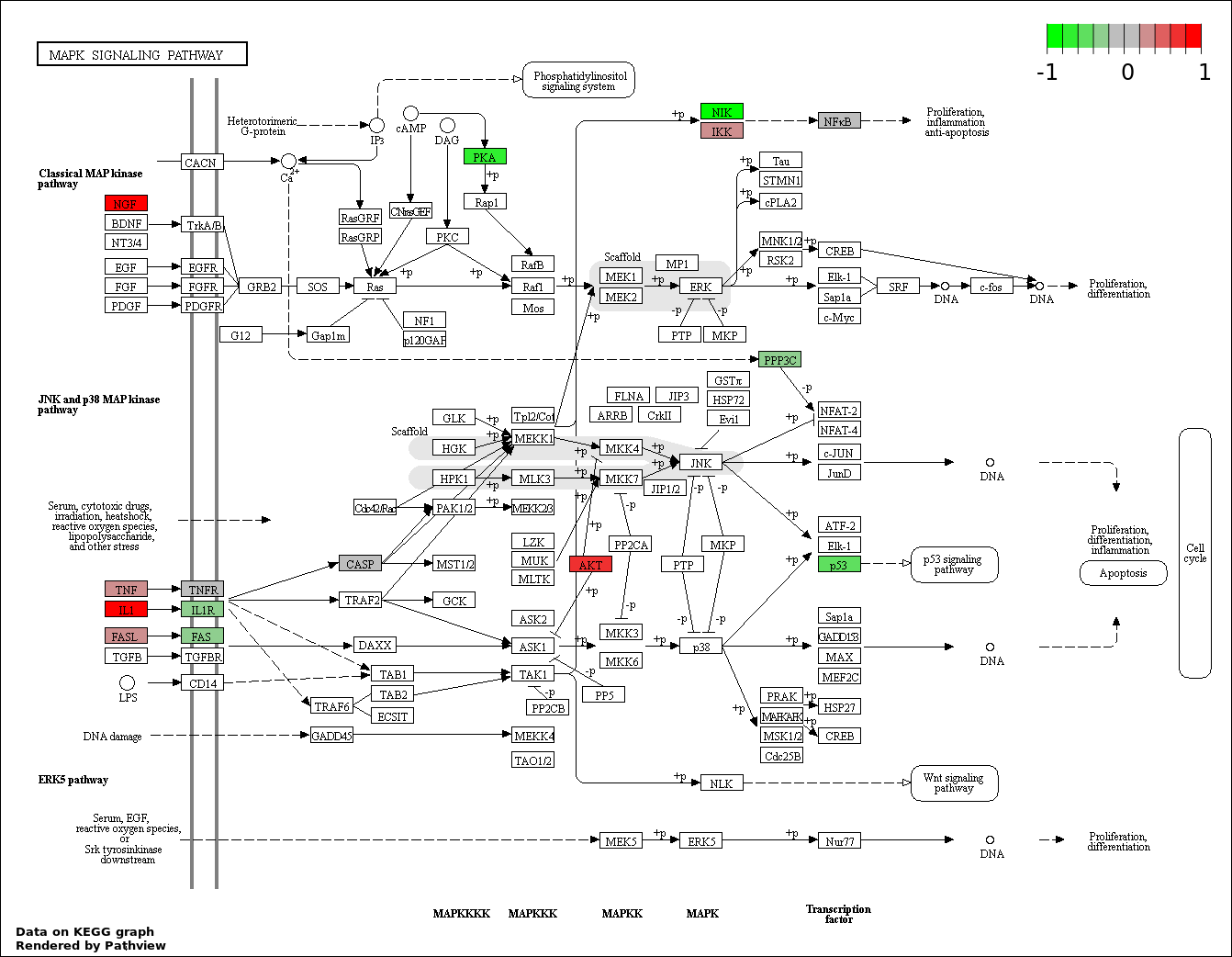

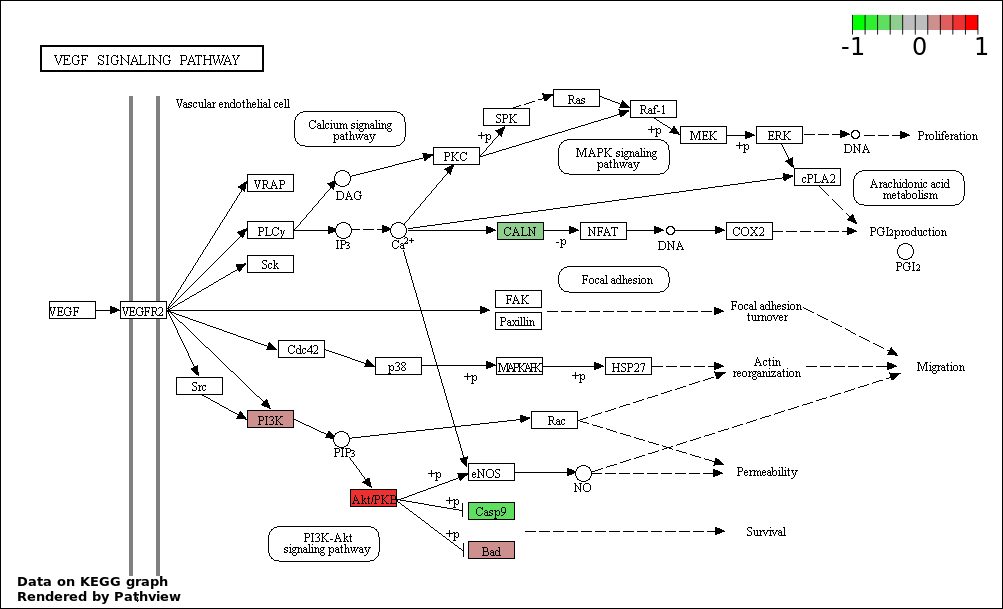









































lncRNA-mediated sponge
| Num | lncRNA | miRNAs | miRNAs count | Gene | Sponge regulatory network | lncRNA log2FC | lncRNA pvalue | Gene log2FC | Gene pvalue | lncRNA-gene Pearson correlation |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | RP11-815I9.4 |
hsa-let-7a-3p;hsa-miR-103a-3p;hsa-miR-146b-5p;hsa-miR-155-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-20a-5p;hsa-miR-26b-5p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-590-5p;hsa-miR-92a-3p | 13 | PRKAR2A | Sponge network | 0.004 | 0.98116 | -1.252 | 0 | 0.731 |
| 2 | RP11-193H5.1 |
hsa-let-7a-3p;hsa-miR-103a-3p;hsa-miR-125a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-197-3p;hsa-miR-20a-5p;hsa-miR-20b-5p;hsa-miR-25-3p;hsa-miR-26b-5p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-7-1-3p | 14 | PRKAR2A | Sponge network | -0.426 | 0.01062 | -1.252 | 0 | 0.655 |
| 3 | RAMP2-AS1 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-365a-3p;hsa-miR-455-5p;hsa-miR-7-5p | 13 | BCL2 | Sponge network | -1.258 | 0.00026 | -0.91 | 1.0E-5 | 0.623 |
| 4 | RP1-193H18.2 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-450b-5p;hsa-miR-452-5p;hsa-miR-542-3p;hsa-miR-7-5p;hsa-miR-96-5p | 13 | BCL2 | Sponge network | -1.539 | 0 | -0.91 | 1.0E-5 | 0.62 |
| 5 | ZNF667-AS1 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-452-5p;hsa-miR-590-5p;hsa-miR-7-5p;hsa-miR-96-5p | 14 | BCL2 | Sponge network | -1.395 | 0 | -0.91 | 1.0E-5 | 0.598 |
| 6 | MAGI2-AS3 |
hsa-miR-103a-3p;hsa-miR-106b-5p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-188-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-29b-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-3607-3p;hsa-miR-424-5p;hsa-miR-429;hsa-miR-532-3p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-629-3p;hsa-miR-93-5p | 30 | PIK3R1 | Sponge network | -0.939 | 4.0E-5 | -1.219 | 0 | 0.589 |
| 7 | RP11-284N8.3 |
hsa-miR-17-5p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-450b-5p;hsa-miR-452-5p;hsa-miR-542-3p;hsa-miR-590-5p;hsa-miR-96-5p | 10 | BCL2 | Sponge network | 0.003 | 0.99478 | -0.91 | 1.0E-5 | 0.579 |
| 8 | BZRAP1-AS1 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-452-5p;hsa-miR-455-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -0.233 | 0.50729 | -0.91 | 1.0E-5 | 0.565 |
| 9 | AC006116.24 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-93-5p | 11 | PIK3R1 | Sponge network | -1.832 | 1.0E-5 | -1.219 | 0 | 0.544 |
| 10 | BZRAP1-AS1 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-584-5p;hsa-miR-93-5p | 12 | PIK3R1 | Sponge network | -0.233 | 0.50729 | -1.219 | 0 | 0.519 |
| 11 | RP11-282O18.3 | hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-29b-3p;hsa-miR-455-3p;hsa-miR-590-5p | 11 | PIK3R1 | Sponge network | -0.266 | 0.05794 | -1.219 | 0 | 0.516 |
| 12 | OIP5-AS1 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-455-5p;hsa-miR-542-3p;hsa-miR-590-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -0.06 | 0.55122 | -0.91 | 1.0E-5 | 0.511 |
| 13 | PWAR6 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-452-5p;hsa-miR-455-5p;hsa-miR-590-5p;hsa-miR-7-5p | 13 | BCL2 | Sponge network | -1.629 | 0 | -0.91 | 1.0E-5 | 0.509 |
| 14 | PCBP1-AS1 | hsa-miR-181b-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-365a-3p;hsa-miR-450b-5p;hsa-miR-452-5p;hsa-miR-455-5p;hsa-miR-542-3p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -0.746 | 0 | -0.91 | 1.0E-5 | 0.509 |
| 15 | RP11-774O3.3 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-365a-3p;hsa-miR-542-3p;hsa-miR-7-5p | 11 | BCL2 | Sponge network | -1.712 | 0 | -0.91 | 1.0E-5 | 0.509 |
| 16 | SNHG14 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-365a-3p;hsa-miR-452-5p;hsa-miR-455-5p;hsa-miR-590-5p;hsa-miR-7-5p | 13 | BCL2 | Sponge network | -1.125 | 0.0001 | -0.91 | 1.0E-5 | 0.505 |
| 17 | MAGI2-AS3 |
hsa-miR-18a-5p;hsa-miR-19b-3p;hsa-miR-205-3p;hsa-miR-21-5p;hsa-miR-30b-5p;hsa-miR-30c-5p;hsa-miR-324-5p;hsa-miR-339-5p;hsa-miR-455-5p;hsa-miR-500a-5p;hsa-miR-576-5p;hsa-miR-92a-3p;hsa-miR-944 | 13 | ATM | Sponge network | -0.939 | 4.0E-5 | -0.093 | 0.45414 | 0.501 |
| 18 | MAGI2-AS3 |
hsa-miR-141-3p;hsa-miR-148b-5p;hsa-miR-188-5p;hsa-miR-200a-3p;hsa-miR-205-5p;hsa-miR-20a-3p;hsa-miR-324-5p;hsa-miR-33a-5p;hsa-miR-429;hsa-miR-452-3p;hsa-miR-501-5p;hsa-miR-589-3p;hsa-miR-660-5p;hsa-miR-708-3p;hsa-miR-92a-3p;hsa-miR-944 | 16 | IRAK3 | Sponge network | -0.939 | 4.0E-5 | -0.224 | 0.27721 | 0.496 |
| 19 | LINC00963 | hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-451a;hsa-miR-452-5p;hsa-miR-455-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -0.285 | 0.0279 | -0.91 | 1.0E-5 | 0.492 |
| 20 | AC093627.10 |
hsa-miR-17-5p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-450b-5p;hsa-miR-455-5p;hsa-miR-590-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -0.893 | 0.00123 | -0.91 | 1.0E-5 | 0.492 |
| 21 | MAGI2-AS3 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-365a-3p;hsa-miR-451a;hsa-miR-452-5p;hsa-miR-455-5p;hsa-miR-7-5p | 14 | BCL2 | Sponge network | -0.939 | 4.0E-5 | -0.91 | 1.0E-5 | 0.488 |
| 22 | FGF14-AS2 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-452-5p;hsa-miR-7-5p;hsa-miR-96-5p | 12 | BCL2 | Sponge network | -1.914 | 0 | -0.91 | 1.0E-5 | 0.488 |
| 23 | RP11-389C8.2 |
hsa-miR-103a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-429;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-93-5p | 12 | PIK3R1 | Sponge network | 0.187 | 0.31051 | -1.219 | 0 | 0.483 |
| 24 | RP11-594N15.3 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-188-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-3607-3p;hsa-miR-590-3p;hsa-miR-93-5p | 23 | PIK3R1 | Sponge network | -2.86 | 0 | -1.219 | 0 | 0.483 |
| 25 | TP73-AS1 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-452-5p;hsa-miR-96-5p | 10 | BCL2 | Sponge network | -0.533 | 0.00643 | -0.91 | 1.0E-5 | 0.483 |
| 26 | AC093627.10 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-590-5p | 13 | PIK3R1 | Sponge network | -0.893 | 0.00123 | -1.219 | 0 | 0.482 |
| 27 | SNHG14 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-584-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p | 15 | PIK3R1 | Sponge network | -1.125 | 0.0001 | -1.219 | 0 | 0.482 |
| 28 | RP11-193H5.1 |
hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-199a-5p;hsa-miR-199b-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-20b-5p;hsa-miR-217;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-320a;hsa-miR-335-3p;hsa-miR-339-5p;hsa-miR-3607-3p | 15 | IL1RAP | Sponge network | -0.426 | 0.01062 | 0.493 | 0.00341 | 0.481 |
| 29 | AC007743.1 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-29b-3p;hsa-miR-320a;hsa-miR-424-5p;hsa-miR-429;hsa-miR-584-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p;hsa-miR-96-5p | 19 | PIK3R1 | Sponge network | -1.053 | 0.00923 | -1.219 | 0 | 0.478 |
| 30 | CTA-221G9.11 |
hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-450b-5p;hsa-miR-629-3p;hsa-miR-93-5p | 12 | PIK3R1 | Sponge network | -2.198 | 0 | -1.219 | 0 | 0.477 |
| 31 | RP1-193H18.2 |
hsa-miR-103a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-455-3p;hsa-miR-584-5p;hsa-miR-589-3p;hsa-miR-96-5p | 15 | PIK3R1 | Sponge network | -1.539 | 0 | -1.219 | 0 | 0.476 |
| 32 | SNHG14 |
hsa-miR-18a-5p;hsa-miR-19b-3p;hsa-miR-205-3p;hsa-miR-21-5p;hsa-miR-324-5p;hsa-miR-339-5p;hsa-miR-455-5p;hsa-miR-576-5p;hsa-miR-590-5p;hsa-miR-92a-3p;hsa-miR-944 | 11 | ATM | Sponge network | -1.125 | 0.0001 | -0.093 | 0.45414 | 0.474 |
| 33 | RP11-473I1.9 |
hsa-miR-146b-5p;hsa-miR-17-5p;hsa-miR-181a-5p;hsa-miR-181b-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-26a-2-3p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-500a-5p;hsa-miR-664a-3p | 13 | XIAP | Sponge network | -0.018 | 0.85711 | -0.014 | 0.8644 | 0.472 |
| 34 | RP11-35G9.5 |
hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-589-3p;hsa-miR-96-5p | 10 | PIK3R1 | Sponge network | -0.33 | 0.08136 | -1.219 | 0 | 0.471 |
| 35 | RP11-400K9.4 |
hsa-miR-103a-3p;hsa-miR-106b-5p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-188-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-324-3p;hsa-miR-93-5p | 12 | PIK3R1 | Sponge network | -0.304 | 0.38627 | -1.219 | 0 | 0.469 |
| 36 | RP4-798P15.3 |
hsa-miR-146b-5p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-29b-1-5p;hsa-miR-320a;hsa-miR-339-5p;hsa-miR-3607-3p;hsa-miR-501-5p;hsa-miR-576-5p;hsa-miR-590-5p | 10 | PIK3CA | Sponge network | -1.213 | 0.00098 | 0.474 | 0.00014 | 0.467 |
| 37 | PWAR6 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p | 16 | PIK3R1 | Sponge network | -1.629 | 0 | -1.219 | 0 | 0.466 |
| 38 | RP11-815I9.4 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-29b-3p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-589-3p;hsa-miR-590-5p | 14 | PIK3R1 | Sponge network | 0.004 | 0.98116 | -1.219 | 0 | 0.464 |
| 39 | RP11-815I9.4 |
hsa-miR-146b-5p;hsa-miR-155-5p;hsa-miR-16-1-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-181a-5p;hsa-miR-181b-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-1-5p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-26a-2-3p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-664a-3p;hsa-miR-92a-3p | 18 | XIAP | Sponge network | 0.004 | 0.98116 | -0.014 | 0.8644 | 0.464 |
| 40 | CTA-221G9.11 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-450b-5p | 11 | BCL2 | Sponge network | -2.198 | 0 | -0.91 | 1.0E-5 | 0.461 |
| 41 | RP4-798P15.3 |
hsa-let-7a-3p;hsa-miR-103a-3p;hsa-miR-146b-5p;hsa-miR-155-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-197-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-26b-5p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-708-3p | 16 | PRKAR2A | Sponge network | -1.213 | 0.00098 | -1.252 | 0 | 0.457 |
| 42 | RP11-284N8.3 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-450b-5p;hsa-miR-455-3p;hsa-miR-590-5p;hsa-miR-93-5p;hsa-miR-96-5p | 14 | PIK3R1 | Sponge network | 0.003 | 0.99478 | -1.219 | 0 | 0.457 |
| 43 | CTD-2015G9.2 |
hsa-miR-103a-3p;hsa-miR-106a-5p;hsa-miR-146b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-197-3p;hsa-miR-20a-5p;hsa-miR-20b-5p;hsa-miR-21-5p;hsa-miR-25-3p;hsa-miR-7-1-3p;hsa-miR-708-3p;hsa-miR-769-3p | 13 | PRKAR2A | Sponge network | -3.112 | 5.0E-5 | -1.252 | 0 | 0.449 |
| 44 | RP11-166D19.1 |
hsa-miR-141-3p;hsa-miR-148b-5p;hsa-miR-188-5p;hsa-miR-200a-3p;hsa-miR-205-5p;hsa-miR-20a-3p;hsa-miR-33a-5p;hsa-miR-429;hsa-miR-452-3p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-708-3p;hsa-miR-92a-3p;hsa-miR-944 | 14 | IRAK3 | Sponge network | -0.882 | 5.0E-5 | -0.224 | 0.27721 | 0.448 |
| 45 | BHLHE40-AS1 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-188-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-450b-5p;hsa-miR-590-5p;hsa-miR-96-5p | 13 | PIK3R1 | Sponge network | -0.932 | 0 | -1.219 | 0 | 0.438 |
| 46 | SH3RF3-AS1 |
hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-188-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-29b-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-590-5p | 16 | PIK3R1 | Sponge network | -0.175 | 0.58985 | -1.219 | 0 | 0.435 |
| 47 | PSMG3-AS1 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-188-5p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-324-3p;hsa-miR-424-5p;hsa-miR-455-3p | 14 | PIK3R1 | Sponge network | -0.793 | 0 | -1.219 | 0 | 0.434 |
| 48 | ZNF667-AS1 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-324-3p;hsa-miR-424-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p;hsa-miR-96-5p | 17 | PIK3R1 | Sponge network | -1.395 | 0 | -1.219 | 0 | 0.434 |
| 49 | FAM66C |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-452-5p;hsa-miR-590-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -0.798 | 0.00038 | -0.91 | 1.0E-5 | 0.43 |
| 50 | AC007743.1 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-365a-3p;hsa-miR-455-5p;hsa-miR-590-5p;hsa-miR-7-5p;hsa-miR-96-5p | 14 | BCL2 | Sponge network | -1.053 | 0.00923 | -0.91 | 1.0E-5 | 0.429 |
| 51 | RAMP2-AS1 |
hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-424-5p;hsa-miR-584-5p;hsa-miR-589-3p | 11 | PIK3R1 | Sponge network | -1.258 | 0.00026 | -1.219 | 0 | 0.429 |
| 52 | RP11-519G16.5 | hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-542-3p;hsa-miR-96-5p | 10 | BCL2 | Sponge network | -2.746 | 2.0E-5 | -0.91 | 1.0E-5 | 0.427 |
| 53 | RP11-693J15.4 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-222-3p;hsa-miR-455-3p;hsa-miR-590-5p | 11 | PIK3R1 | Sponge network | -0.732 | 0.18487 | -1.219 | 0 | 0.426 |
| 54 | NOP14-AS1 |
hsa-miR-127-3p;hsa-miR-129-5p;hsa-miR-145-5p;hsa-miR-148a-3p;hsa-miR-26a-2-3p;hsa-miR-26a-5p;hsa-miR-30a-3p;hsa-miR-30e-3p;hsa-miR-628-5p;hsa-miR-664a-3p | 10 | DFFA | Sponge network | 0.939 | 0 | 0.106 | 0.16666 | 0.422 |
| 55 | OIP5-AS1 |
hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-29b-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-590-5p | 11 | PIK3R1 | Sponge network | -0.06 | 0.55122 | -1.219 | 0 | 0.421 |
| 56 | OIP5-AS1 |
hsa-miR-146b-5p;hsa-miR-17-5p;hsa-miR-181a-2-3p;hsa-miR-181b-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-29a-5p;hsa-miR-500a-5p | 10 | XIAP | Sponge network | -0.06 | 0.55122 | -0.014 | 0.8644 | 0.419 |
| 57 | RP11-774O3.3 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-188-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-324-3p;hsa-miR-424-5p;hsa-miR-589-3p;hsa-miR-629-3p | 19 | PIK3R1 | Sponge network | -1.712 | 0 | -1.219 | 0 | 0.415 |
| 58 | RP4-798P15.3 |
hsa-miR-18a-5p;hsa-miR-19b-3p;hsa-miR-205-3p;hsa-miR-21-5p;hsa-miR-30c-5p;hsa-miR-324-5p;hsa-miR-339-5p;hsa-miR-455-5p;hsa-miR-500a-5p;hsa-miR-576-5p;hsa-miR-590-5p;hsa-miR-944 | 12 | ATM | Sponge network | -1.213 | 0.00098 | -0.093 | 0.45414 | 0.411 |
| 59 | TPTEP1 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-365a-3p;hsa-miR-455-5p;hsa-miR-542-3p;hsa-miR-590-5p;hsa-miR-7-5p | 14 | BCL2 | Sponge network | -2.193 | 0 | -0.91 | 1.0E-5 | 0.409 |
| 60 | RP11-359B12.2 | hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p | 10 | PIK3R1 | Sponge network | -0.334 | 0.03398 | -1.219 | 0 | 0.409 |
| 61 | FGF14-AS2 |
hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-629-3p;hsa-miR-96-5p | 12 | PIK3R1 | Sponge network | -1.914 | 0 | -1.219 | 0 | 0.405 |
| 62 | RP11-13K12.5 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-29a-5p;hsa-miR-365a-3p;hsa-miR-451a;hsa-miR-7-5p | 11 | BCL2 | Sponge network | -0.552 | 0.43847 | -0.91 | 1.0E-5 | 0.403 |
| 63 | RP11-58E21.3 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-140-5p;hsa-miR-190a-5p;hsa-miR-192-5p;hsa-miR-199a-5p;hsa-miR-199b-5p;hsa-miR-20b-5p;hsa-miR-217;hsa-miR-335-3p;hsa-miR-93-5p | 11 | IL1RAP | Sponge network | 0.445 | 0.04375 | 0.493 | 0.00341 | 0.402 |
| 64 | WDR86-AS1 | hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-188-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-324-3p;hsa-miR-455-3p;hsa-miR-93-5p;hsa-miR-96-5p | 11 | PIK3R1 | Sponge network | -0.396 | 0.35778 | -1.219 | 0 | 0.402 |
| 65 | RP11-166D19.1 |
hsa-miR-103a-3p;hsa-miR-106b-5p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-188-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-320b;hsa-miR-324-3p;hsa-miR-3607-3p;hsa-miR-429;hsa-miR-455-3p;hsa-miR-532-3p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-96-5p | 28 | PIK3R1 | Sponge network | -0.882 | 5.0E-5 | -1.219 | 0 | 0.401 |
| 66 | RP11-594N15.3 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -2.86 | 0 | -0.91 | 1.0E-5 | 0.4 |
| 67 | CTD-2015G9.2 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-450b-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-93-5p | 18 | PIK3R1 | Sponge network | -3.112 | 5.0E-5 | -1.219 | 0 | 0.397 |
| 68 | RASSF8-AS1 |
hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-324-3p;hsa-miR-429;hsa-miR-629-3p;hsa-miR-93-5p | 13 | PIK3R1 | Sponge network | 0.121 | 0.52445 | -1.219 | 0 | 0.395 |
| 69 | RP11-400K9.4 |
hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-339-5p;hsa-miR-362-5p;hsa-miR-93-5p | 14 | PRKACB | Sponge network | -0.304 | 0.38627 | -0.321 | 0.02288 | 0.393 |
| 70 | RP4-798P15.3 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-29b-3p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-96-5p | 19 | PIK3R1 | Sponge network | -1.213 | 0.00098 | -1.219 | 0 | 0.393 |
| 71 | LINC00883 | hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-30d-3p;hsa-miR-362-3p;hsa-miR-532-5p;hsa-miR-944 | 14 | PRKACB | Sponge network | 0.556 | 0.00093 | -0.321 | 0.02288 | 0.392 |
| 72 | RP11-193H5.1 |
hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-1-5p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-25-3p;hsa-miR-26a-2-3p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-500a-5p | 13 | XIAP | Sponge network | -0.426 | 0.01062 | -0.014 | 0.8644 | 0.391 |
| 73 | MAGI2-AS3 |
hsa-let-7a-3p;hsa-let-7d-3p;hsa-miR-141-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-194-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-30d-3p;hsa-miR-339-5p;hsa-miR-362-5p;hsa-miR-429;hsa-miR-582-5p;hsa-miR-7-1-3p;hsa-miR-708-3p;hsa-miR-93-5p;hsa-miR-944 | 24 | PRKACB | Sponge network | -0.939 | 4.0E-5 | -0.321 | 0.02288 | 0.386 |
| 74 | TP73-AS1 |
hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-96-5p | 13 | PIK3R1 | Sponge network | -0.533 | 0.00643 | -1.219 | 0 | 0.386 |
| 75 | MIR22HG |
hsa-miR-130b-3p;hsa-miR-16-1-3p;hsa-miR-181c-5p;hsa-miR-181d-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200b-3p;hsa-miR-205-3p;hsa-miR-301a-3p;hsa-miR-335-3p | 10 | ENDOD1 | Sponge network | -0.601 | 4.0E-5 | -0.554 | 0.00014 | 0.384 |
| 76 | CTD-2135D7.5 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-185-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p | 12 | PIK3R1 | Sponge network | -2.579 | 0.0001 | -1.219 | 0 | 0.379 |
| 77 | RP11-594N15.3 |
hsa-let-7a-3p;hsa-let-7d-3p;hsa-miR-141-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-194-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-582-5p;hsa-miR-93-5p | 17 | PRKACB | Sponge network | -2.86 | 0 | -0.321 | 0.02288 | 0.377 |
| 78 | LINC00900 |
hsa-miR-103a-3p;hsa-miR-106b-5p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-188-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-3607-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-93-5p | 26 | PIK3R1 | Sponge network | -1.803 | 0 | -1.219 | 0 | 0.373 |
| 79 | DLGAP1-AS5 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-450b-5p;hsa-miR-455-5p;hsa-miR-96-5p | 11 | BCL2 | Sponge network | -6.207 | 0 | -0.91 | 1.0E-5 | 0.373 |
| 80 | AC003090.1 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-424-5p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p;hsa-miR-96-5p | 23 | PIK3R1 | Sponge network | -4.323 | 0 | -1.219 | 0 | 0.369 |
| 81 | LINC00957 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-450b-5p;hsa-miR-455-5p | 11 | BCL2 | Sponge network | -0.677 | 2.0E-5 | -0.91 | 1.0E-5 | 0.368 |
| 82 | RP11-757G1.6 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-424-5p;hsa-miR-532-3p;hsa-miR-589-3p;hsa-miR-93-5p | 15 | PIK3R1 | Sponge network | -1.346 | 0.00088 | -1.219 | 0 | 0.367 |
| 83 | RP11-166D19.1 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-452-5p;hsa-miR-590-5p;hsa-miR-96-5p | 10 | BCL2 | Sponge network | -0.882 | 5.0E-5 | -0.91 | 1.0E-5 | 0.367 |
| 84 | RP11-517P14.2 | hsa-miR-16-1-3p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-1-5p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-29a-5p;hsa-miR-3607-3p;hsa-miR-500a-5p | 10 | XIAP | Sponge network | -0.402 | 0.1503 | -0.014 | 0.8644 | 0.364 |
| 85 | FAM225B |
hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-30d-3p;hsa-miR-34a-5p;hsa-miR-362-3p;hsa-miR-362-5p;hsa-miR-582-5p | 13 | PRKACB | Sponge network | 2.186 | 0 | -0.321 | 0.02288 | 0.363 |
| 86 | RP11-473I1.9 |
hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-199a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-320a;hsa-miR-339-5p;hsa-miR-454-3p;hsa-miR-590-5p;hsa-miR-664a-3p | 13 | IL1RAP | Sponge network | -0.018 | 0.85711 | 0.493 | 0.00341 | 0.358 |
| 87 | LINC00639 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-629-3p;hsa-miR-93-5p;hsa-miR-96-5p | 14 | PIK3R1 | Sponge network | -0.91 | 0.00848 | -1.219 | 0 | 0.354 |
| 88 | EMX2OS |
hsa-miR-103a-3p;hsa-miR-106b-5p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-188-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-29b-3p;hsa-miR-324-3p;hsa-miR-3607-3p;hsa-miR-532-3p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-629-3p;hsa-miR-93-5p | 27 | PIK3R1 | Sponge network | -1.459 | 1.0E-5 | -1.219 | 0 | 0.349 |
| 89 | CTC-205M6.5 |
hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-30d-3p;hsa-miR-339-5p;hsa-miR-34a-5p;hsa-miR-7-1-3p;hsa-miR-944;hsa-miR-96-5p | 16 | PRKACB | Sponge network | 0.381 | 0.01756 | -0.321 | 0.02288 | 0.348 |
| 90 | RP4-798P15.3 |
hsa-miR-146b-5p;hsa-miR-155-5p;hsa-miR-16-1-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-181a-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-1-5p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-500a-5p;hsa-miR-708-3p | 16 | XIAP | Sponge network | -1.213 | 0.00098 | -0.014 | 0.8644 | 0.347 |
| 91 | AC003090.1 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-365a-3p;hsa-miR-452-5p;hsa-miR-590-5p;hsa-miR-96-5p | 11 | BCL2 | Sponge network | -4.323 | 0 | -0.91 | 1.0E-5 | 0.347 |
| 92 | RP11-774O3.3 |
hsa-miR-141-3p;hsa-miR-148b-5p;hsa-miR-188-5p;hsa-miR-205-5p;hsa-miR-20a-3p;hsa-miR-324-5p;hsa-miR-33a-5p;hsa-miR-452-3p;hsa-miR-589-3p;hsa-miR-944 | 10 | IRAK3 | Sponge network | -1.712 | 0 | -0.224 | 0.27721 | 0.346 |
| 93 | AC011738.4 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-140-5p;hsa-miR-186-5p;hsa-miR-192-5p;hsa-miR-19b-3p;hsa-miR-20b-5p;hsa-miR-217;hsa-miR-29b-3p;hsa-miR-29c-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-3607-3p;hsa-miR-361-5p;hsa-miR-664a-3p | 15 | IL1RAP | Sponge network | 3.153 | 0 | 0.493 | 0.00341 | 0.345 |
| 94 | CTD-2008P7.9 |
hsa-let-7a-3p;hsa-miR-103a-3p;hsa-miR-125a-5p;hsa-miR-146b-5p;hsa-miR-155-5p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-197-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-25-3p;hsa-miR-29a-5p;hsa-miR-590-5p | 13 | PRKAR2A | Sponge network | -1.202 | 0.00093 | -1.252 | 0 | 0.344 |
| 95 | MIR497HG |
hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-324-3p;hsa-miR-455-3p;hsa-miR-584-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p;hsa-miR-96-5p | 20 | PIK3R1 | Sponge network | -1.263 | 0.00248 | -1.219 | 0 | 0.343 |
| 96 | EMX2OS |
hsa-miR-130b-5p;hsa-miR-17-3p;hsa-miR-186-5p;hsa-miR-192-5p;hsa-miR-197-3p;hsa-miR-26b-5p;hsa-miR-30d-3p;hsa-miR-335-3p;hsa-miR-590-3p;hsa-miR-944 | 10 | IL1R1 | Sponge network | -1.459 | 1.0E-5 | -0.258 | 0.08776 | 0.342 |
| 97 | MIR497HG |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-590-5p;hsa-miR-7-5p;hsa-miR-96-5p | 10 | BCL2 | Sponge network | -1.263 | 0.00248 | -0.91 | 1.0E-5 | 0.342 |
| 98 | SNHG14 |
hsa-miR-141-3p;hsa-miR-148b-5p;hsa-miR-205-5p;hsa-miR-20a-3p;hsa-miR-324-5p;hsa-miR-452-3p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-708-3p;hsa-miR-92a-3p;hsa-miR-944 | 11 | IRAK3 | Sponge network | -1.125 | 0.0001 | -0.224 | 0.27721 | 0.34 |
| 99 | RP11-403I13.7 | hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-339-5p;hsa-miR-362-5p;hsa-miR-532-5p;hsa-miR-590-5p;hsa-miR-96-5p | 14 | PRKACB | Sponge network | 0.214 | 0.38819 | -0.321 | 0.02288 | 0.34 |
| 100 | RASSF8-AS1 |
hsa-miR-141-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-429;hsa-miR-93-5p;hsa-miR-944 | 14 | PRKACB | Sponge network | 0.121 | 0.52445 | -0.321 | 0.02288 | 0.338 |
| 101 | MAGI2-AS3 |
hsa-miR-141-3p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-200b-3p;hsa-miR-25-3p;hsa-miR-30b-5p;hsa-miR-30c-5p;hsa-miR-320a;hsa-miR-320b;hsa-miR-342-3p;hsa-miR-429;hsa-miR-590-3p;hsa-miR-944 | 15 | PRKAR1A | Sponge network | -0.939 | 4.0E-5 | -0.39 | 0 | 0.334 |
| 102 | LINC00284 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p | 19 | PIK3R1 | Sponge network | -4.159 | 0 | -1.219 | 0 | 0.333 |
| 103 | RP11-557H15.3 | hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-140-5p;hsa-miR-186-5p;hsa-miR-190a-5p;hsa-miR-20b-5p;hsa-miR-217;hsa-miR-29c-3p;hsa-miR-335-3p;hsa-miR-361-5p;hsa-miR-664a-3p | 11 | IL1RAP | Sponge network | 1.673 | 8.0E-5 | 0.493 | 0.00341 | 0.332 |
| 104 | DHRS4-AS1 | hsa-miR-146b-5p;hsa-miR-16-1-3p;hsa-miR-17-5p;hsa-miR-181a-5p;hsa-miR-181b-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-500a-5p | 10 | XIAP | Sponge network | -0.068 | 0.72321 | -0.014 | 0.8644 | 0.33 |
| 105 | LINC00284 |
hsa-miR-148b-5p;hsa-miR-205-5p;hsa-miR-20a-3p;hsa-miR-324-5p;hsa-miR-33a-5p;hsa-miR-452-3p;hsa-miR-501-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-944 | 10 | IRAK3 | Sponge network | -4.159 | 0 | -0.224 | 0.27721 | 0.328 |
| 106 | DNM3OS |
hsa-miR-106b-5p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-224-3p;hsa-miR-29b-3p;hsa-miR-429;hsa-miR-590-5p;hsa-miR-93-5p;hsa-miR-96-5p | 16 | PIK3R1 | Sponge network | 0.932 | 0.00442 | -1.219 | 0 | 0.328 |
| 107 | RP11-362F19.1 | hsa-miR-17-5p;hsa-miR-188-5p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-424-5p;hsa-miR-584-5p;hsa-miR-589-3p | 10 | PIK3R1 | Sponge network | -0.803 | 0.19311 | -1.219 | 0 | 0.328 |
| 108 | RP11-68I18.10 |
hsa-miR-103a-3p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p | 12 | PIK3R1 | Sponge network | -1.464 | 0.00126 | -1.219 | 0 | 0.327 |
| 109 | HHIP-AS1 |
hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-29b-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-532-3p;hsa-miR-590-3p | 14 | PIK3R1 | Sponge network | -1.293 | 0.01561 | -1.219 | 0 | 0.327 |
| 110 | BHLHE40-AS1 |
hsa-let-7a-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-339-5p;hsa-miR-362-5p;hsa-miR-590-5p;hsa-miR-96-5p | 12 | PRKACB | Sponge network | -0.932 | 0 | -0.321 | 0.02288 | 0.322 |
| 111 | RP11-757G1.6 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-365a-3p;hsa-miR-452-5p | 10 | BCL2 | Sponge network | -1.346 | 0.00088 | -0.91 | 1.0E-5 | 0.317 |
| 112 | LINC00284 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-365a-3p;hsa-miR-455-5p;hsa-miR-542-3p;hsa-miR-590-5p;hsa-miR-7-5p | 13 | BCL2 | Sponge network | -4.159 | 0 | -0.91 | 1.0E-5 | 0.316 |
| 113 | AF127936.7 |
hsa-miR-155-5p;hsa-miR-16-1-3p;hsa-miR-16-2-3p;hsa-miR-19b-1-5p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-25-3p;hsa-miR-26a-2-3p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-500a-5p;hsa-miR-664a-3p | 12 | XIAP | Sponge network | 0.453 | 0.00811 | -0.014 | 0.8644 | 0.315 |
| 114 | HHIP-AS1 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-451a;hsa-miR-455-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -1.293 | 0.01561 | -0.91 | 1.0E-5 | 0.315 |
| 115 | DLGAP1-AS5 |
hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-589-3p;hsa-miR-96-5p | 13 | PIK3R1 | Sponge network | -6.207 | 0 | -1.219 | 0 | 0.315 |
| 116 | MAGI2-AS3 |
hsa-let-7a-3p;hsa-miR-103a-3p;hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-197-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-25-3p;hsa-miR-26b-5p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-320b;hsa-miR-3607-3p;hsa-miR-582-5p;hsa-miR-7-1-3p;hsa-miR-708-3p;hsa-miR-769-3p;hsa-miR-92a-3p | 21 | PRKAR2A | Sponge network | -0.939 | 4.0E-5 | -1.252 | 0 | 0.314 |
| 117 | TPTEP1 |
hsa-miR-18a-5p;hsa-miR-19b-3p;hsa-miR-205-3p;hsa-miR-21-5p;hsa-miR-324-5p;hsa-miR-339-5p;hsa-miR-455-5p;hsa-miR-576-5p;hsa-miR-590-5p;hsa-miR-92a-3p;hsa-miR-944 | 11 | ATM | Sponge network | -2.193 | 0 | -0.093 | 0.45414 | 0.313 |
| 118 | AF127936.7 |
hsa-miR-106a-5p;hsa-miR-140-5p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-20b-5p;hsa-miR-217;hsa-miR-320a;hsa-miR-339-5p;hsa-miR-3607-3p;hsa-miR-362-3p;hsa-miR-454-3p;hsa-miR-664a-3p | 12 | IL1RAP | Sponge network | 0.453 | 0.00811 | 0.493 | 0.00341 | 0.313 |
| 119 | RP11-35G9.5 |
hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-96-5p | 13 | PRKACB | Sponge network | -0.33 | 0.08136 | -0.321 | 0.02288 | 0.312 |
| 120 | RP11-815I9.4 |
hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-199b-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-29b-3p;hsa-miR-301a-3p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-590-5p;hsa-miR-664a-3p | 12 | IL1RAP | Sponge network | 0.004 | 0.98116 | 0.493 | 0.00341 | 0.308 |
| 121 | TPTEP1 |
hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-3607-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p | 20 | PIK3R1 | Sponge network | -2.193 | 0 | -1.219 | 0 | 0.308 |
| 122 | LINC00957 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-455-3p | 14 | PIK3R1 | Sponge network | -0.677 | 2.0E-5 | -1.219 | 0 | 0.306 |
| 123 | PSMG3-AS1 |
hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200c-3p;hsa-miR-339-5p | 12 | PRKACB | Sponge network | -0.793 | 0 | -0.321 | 0.02288 | 0.303 |
| 124 | RP11-161M6.2 | hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-324-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-589-3p | 11 | PIK3R1 | Sponge network | -0.977 | 0.00013 | -1.219 | 0 | 0.299 |
| 125 | BZRAP1-AS1 |
hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-339-5p;hsa-miR-93-5p;hsa-miR-944 | 13 | PRKACB | Sponge network | -0.233 | 0.50729 | -0.321 | 0.02288 | 0.299 |
| 126 | RP11-193H5.1 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-29b-3p;hsa-miR-320a;hsa-miR-3607-3p | 12 | PIK3R1 | Sponge network | -0.426 | 0.01062 | -1.219 | 0 | 0.297 |
| 127 | RP11-384L8.1 |
hsa-miR-17-5p;hsa-miR-181b-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-365a-3p;hsa-miR-7-5p | 11 | BCL2 | Sponge network | -1.152 | 0.00016 | -0.91 | 1.0E-5 | 0.296 |
| 128 | AC007743.1 |
hsa-let-7a-3p;hsa-miR-146b-5p;hsa-miR-155-5p;hsa-miR-17-5p;hsa-miR-197-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-342-5p;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-92a-3p | 13 | PRKAR2A | Sponge network | -1.053 | 0.00923 | -1.252 | 0 | 0.295 |
| 129 | CTD-2135D7.5 |
hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-21-5p;hsa-miR-224-3p;hsa-miR-24-2-5p;hsa-miR-450b-5p;hsa-miR-452-5p;hsa-miR-455-5p;hsa-miR-542-3p;hsa-miR-590-5p;hsa-miR-7-5p | 11 | BCL2 | Sponge network | -2.579 | 0.0001 | -0.91 | 1.0E-5 | 0.294 |
| 130 | LINC00702 |
hsa-miR-103a-3p;hsa-miR-106b-5p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-186-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-29b-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-324-3p;hsa-miR-532-3p;hsa-miR-589-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-629-3p;hsa-miR-93-5p;hsa-miR-96-5p | 26 | PIK3R1 | Sponge network | -0.573 | 0.0699 | -1.219 | 0 | 0.294 |
| 131 | RP11-327P2.5 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-188-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-590-5p | 11 | PIK3R1 | Sponge network | -0.574 | 2.0E-5 | -1.219 | 0 | 0.292 |
| 132 | SH3RF3-AS1 |
hsa-let-7a-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-26b-5p;hsa-miR-29a-5p;hsa-miR-320a;hsa-miR-320b;hsa-miR-342-5p;hsa-miR-582-5p;hsa-miR-590-5p;hsa-miR-7-1-3p | 11 | PRKAR2A | Sponge network | -0.175 | 0.58985 | -1.252 | 0 | 0.291 |
| 133 | MAGI2-AS3 |
hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-5p;hsa-miR-20a-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-339-5p;hsa-miR-361-5p;hsa-miR-421;hsa-miR-429;hsa-miR-452-5p;hsa-miR-7-1-3p | 12 | PPP3CB | Sponge network | -0.939 | 4.0E-5 | -0.431 | 0 | 0.29 |
| 134 | RP11-434B12.1 | hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-320b;hsa-miR-429;hsa-miR-629-3p | 10 | PIK3R1 | Sponge network | -0.525 | 0.01648 | -1.219 | 0 | 0.289 |
| 135 | LINC00865 |
hsa-miR-15a-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-320a;hsa-miR-3607-3p;hsa-miR-424-5p;hsa-miR-629-3p | 13 | PIK3R1 | Sponge network | -0.336 | 0.22355 | -1.219 | 0 | 0.288 |
| 136 | RP11-284N8.3 |
hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200c-3p;hsa-miR-590-5p;hsa-miR-93-5p;hsa-miR-944;hsa-miR-96-5p | 14 | PRKACB | Sponge network | 0.003 | 0.99478 | -0.321 | 0.02288 | 0.281 |
| 137 | LINC00900 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-365a-3p;hsa-miR-455-5p;hsa-miR-590-5p;hsa-miR-7-5p | 11 | BCL2 | Sponge network | -1.803 | 0 | -0.91 | 1.0E-5 | 0.281 |
| 138 | RP11-166D19.1 |
hsa-let-7a-3p;hsa-let-7d-3p;hsa-miR-141-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-30d-3p;hsa-miR-339-5p;hsa-miR-429;hsa-miR-590-5p;hsa-miR-7-1-3p;hsa-miR-708-3p;hsa-miR-944;hsa-miR-96-5p | 22 | PRKACB | Sponge network | -0.882 | 5.0E-5 | -0.321 | 0.02288 | 0.279 |
| 139 | RP11-244O19.1 |
hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-30d-3p;hsa-miR-34a-5p;hsa-miR-362-5p;hsa-miR-532-5p;hsa-miR-93-5p | 13 | PRKACB | Sponge network | 0.52 | 0.05156 | -0.321 | 0.02288 | 0.278 |
| 140 | CTD-2554C21.3 |
hsa-miR-17-5p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-224-5p;hsa-miR-24-2-5p;hsa-miR-29a-5p;hsa-miR-455-5p;hsa-miR-7-5p | 10 | BCL2 | Sponge network | -2.118 | 6.0E-5 | -0.91 | 1.0E-5 | 0.277 |
| 141 | RP11-321G12.1 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-589-3p | 11 | PIK3R1 | Sponge network | -3.552 | 0 | -1.219 | 0 | 0.273 |
| 142 | RP11-399O19.9 | hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-429;hsa-miR-93-5p;hsa-miR-944 | 11 | PRKACB | Sponge network | -0.216 | 0.29068 | -0.321 | 0.02288 | 0.273 |
| 143 | LINC00900 |
hsa-miR-141-3p;hsa-miR-148b-5p;hsa-miR-188-5p;hsa-miR-200a-3p;hsa-miR-205-5p;hsa-miR-20a-3p;hsa-miR-324-5p;hsa-miR-33a-5p;hsa-miR-452-3p;hsa-miR-501-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-708-3p;hsa-miR-92a-3p | 14 | IRAK3 | Sponge network | -1.803 | 0 | -0.224 | 0.27721 | 0.27 |
| 144 | RP11-411K7.1 |
hsa-miR-125a-5p;hsa-miR-130a-3p;hsa-miR-130b-3p;hsa-miR-15b-5p;hsa-miR-301a-3p;hsa-miR-320a;hsa-miR-320b;hsa-miR-34c-5p;hsa-miR-421;hsa-miR-550a-5p | 10 | CFLAR | Sponge network | -0.883 | 0.003 | -0.347 | 0.00043 | 0.268 |
| 145 | CTC-297N7.7 |
hsa-miR-103a-3p;hsa-miR-1301-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-20a-5p;hsa-miR-222-3p;hsa-miR-224-3p;hsa-miR-455-3p;hsa-miR-589-3p;hsa-miR-629-3p;hsa-miR-96-5p | 16 | PIK3R1 | Sponge network | -1.666 | 0.01558 | -1.219 | 0 | 0.267 |
| 146 | CTD-2554C21.3 |
hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-186-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-455-3p;hsa-miR-629-3p | 13 | PIK3R1 | Sponge network | -2.118 | 6.0E-5 | -1.219 | 0 | 0.267 |
| 147 | RP11-389C8.2 |
hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-429;hsa-miR-93-5p;hsa-miR-944 | 15 | PRKACB | Sponge network | 0.187 | 0.31051 | -0.321 | 0.02288 | 0.267 |
| 148 | RP11-815I9.4 |
hsa-let-7a-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-34a-5p;hsa-miR-590-5p | 10 | PRKACB | Sponge network | 0.004 | 0.98116 | -0.321 | 0.02288 | 0.266 |
| 149 | FGD5-AS1 | hsa-let-7a-3p;hsa-miR-141-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-362-5p;hsa-miR-944 | 11 | PRKACB | Sponge network | -0.292 | 0.01482 | -0.321 | 0.02288 | 0.265 |
| 150 | RP11-720L2.4 | hsa-let-7a-3p;hsa-let-7d-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-93-5p | 11 | PRKACB | Sponge network | -0.317 | 0.39052 | -0.321 | 0.02288 | 0.265 |
| 151 | RP11-400K9.4 |
hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-17-5p;hsa-miR-186-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-5p;hsa-miR-301a-3p;hsa-miR-339-5p;hsa-miR-93-5p | 10 | IL1RAP | Sponge network | -0.304 | 0.38627 | 0.493 | 0.00341 | 0.262 |
| 152 | AC006116.24 |
hsa-let-7a-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200c-3p;hsa-miR-93-5p | 10 | PRKACB | Sponge network | -1.832 | 1.0E-5 | -0.321 | 0.02288 | 0.261 |
| 153 | RP11-296E23.1 | hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-2-3p;hsa-miR-16-5p;hsa-miR-182-5p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-21-5p;hsa-miR-224-3p | 10 | PIK3R1 | Sponge network | -4.595 | 0 | -1.219 | 0 | 0.261 |
| 154 | FAM66C |
hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-22-5p;hsa-miR-221-3p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-455-3p;hsa-miR-590-5p | 10 | PIK3R1 | Sponge network | -0.798 | 0.00038 | -1.219 | 0 | 0.26 |
| 155 | RP11-400K9.4 |
hsa-let-7a-3p;hsa-miR-103a-3p;hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-197-3p;hsa-miR-20a-5p;hsa-miR-25-3p;hsa-miR-26b-5p | 10 | PRKAR2A | Sponge network | -0.304 | 0.38627 | -1.252 | 0 | 0.26 |
| 156 | FLG-AS1 |
hsa-miR-106b-5p;hsa-miR-146b-5p;hsa-miR-155-5p;hsa-miR-16-5p;hsa-miR-197-3p;hsa-miR-21-5p;hsa-miR-25-3p;hsa-miR-320b;hsa-miR-342-5p;hsa-miR-3913-5p;hsa-miR-7-1-3p | 11 | PRKAR2A | Sponge network | -1.901 | 0 | -1.252 | 0 | 0.258 |
| 157 | RP4-798P15.3 |
hsa-miR-141-3p;hsa-miR-148b-5p;hsa-miR-205-5p;hsa-miR-20a-3p;hsa-miR-324-5p;hsa-miR-452-3p;hsa-miR-501-5p;hsa-miR-589-3p;hsa-miR-590-5p;hsa-miR-660-5p;hsa-miR-708-3p;hsa-miR-944 | 12 | IRAK3 | Sponge network | -1.213 | 0.00098 | -0.224 | 0.27721 | 0.258 |
| 158 | SLC26A4-AS1 | hsa-miR-103a-3p;hsa-miR-15a-5p;hsa-miR-17-5p;hsa-miR-185-5p;hsa-miR-20a-5p;hsa-miR-22-5p;hsa-miR-222-3p;hsa-miR-424-5p;hsa-miR-450b-5p;hsa-miR-589-3p | 10 | PIK3R1 | Sponge network | -1.114 | 0.08689 | -1.219 | 0 | 0.257 |
| 159 | RP11-58E21.3 |
hsa-miR-103a-3p;hsa-miR-106a-5p;hsa-miR-106b-5p;hsa-miR-125a-5p;hsa-miR-182-5p;hsa-miR-20b-5p;hsa-miR-25-3p;hsa-miR-26b-5p;hsa-miR-3913-5p;hsa-miR-582-5p;hsa-miR-708-3p;hsa-miR-769-3p | 12 | PRKAR2A | Sponge network | 0.445 | 0.04375 | -1.252 | 0 | 0.255 |
| 160 | RP4-794H19.1 |
hsa-miR-141-3p;hsa-miR-16-2-3p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-194-3p;hsa-miR-19b-3p;hsa-miR-200a-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-30d-3p;hsa-miR-362-5p;hsa-miR-582-5p | 13 | PRKACB | Sponge network | 1.019 | 8.0E-5 | -0.321 | 0.02288 | 0.255 |
| 161 | RP11-456K23.1 |
hsa-miR-130b-3p;hsa-miR-16-1-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200b-3p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-26b-3p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-3607-3p;hsa-miR-3613-5p;hsa-miR-362-3p;hsa-miR-429;hsa-miR-454-3p;hsa-miR-589-3p;hsa-miR-590-3p | 17 | ENDOD1 | Sponge network | -0.223 | 0.27461 | -0.554 | 0.00014 | 0.254 |
| 162 | SH3RF3-AS1 |
hsa-let-7a-3p;hsa-miR-16-2-3p;hsa-miR-182-5p;hsa-miR-183-5p;hsa-miR-18a-5p;hsa-miR-193a-3p;hsa-miR-194-3p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200b-3p;hsa-miR-200c-3p;hsa-miR-339-5p;hsa-miR-582-5p;hsa-miR-590-5p;hsa-miR-7-1-3p | 15 | PRKACB | Sponge network | -0.175 | 0.58985 | -0.321 | 0.02288 | 0.254 |
| 163 | RP5-1042I8.7 | hsa-miR-103a-3p;hsa-miR-106b-5p;hsa-miR-1301-3p;hsa-miR-15b-5p;hsa-miR-17-5p;hsa-miR-182-5p;hsa-miR-20a-5p;hsa-miR-21-5p;hsa-miR-93-5p;hsa-miR-96-5p | 10 | PIK3R1 | Sponge network | -0.734 | 2.0E-5 | -1.219 | 0 | 0.252 |
| 164 | LINC00702 |
hsa-miR-130b-3p;hsa-miR-16-1-3p;hsa-miR-192-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-200b-3p;hsa-miR-205-3p;hsa-miR-20a-3p;hsa-miR-26b-3p;hsa-miR-301a-3p;hsa-miR-335-3p;hsa-miR-361-5p;hsa-miR-454-3p;hsa-miR-589-3p;hsa-miR-590-3p | 15 | ENDOD1 | Sponge network | -0.573 | 0.0699 | -0.554 | 0.00014 | 0.252 |