This regulatory network was inferred from the input dataset. The miRNAs and mRNAs are
presented as round and rectangle nodes respectively. The numerical value popped up upon mouse over the gene node is the log2 transformed fold-change of the gene expression between the two groups. All of the nodes are clickable, and the detailed information of the miRNAs/mRNAs and related cancer pathway will be displayed in another window. The edges between nodes are supported by both interactions (predicted or experimentally verified) and correlations learnt from cancer dataset. The numerical value popped up upon mouse over the edge is the correlation beat value (effect size) between the two nodes. The experimental evidences of the edges reported in previous cancer studies are highlighted by red/orange color. All of these information can be accessed by the "mouse-over" action. This network shows a full map of the miRNA-mRNA regulation of the input gene list(s), and the hub miRNAs (with the high network degree/betweenness centrality) would be the potential cancer drivers or tumor suppressors. The full result table can be accessed in the "Regulations" tab.
"miRNACancerMAP" is also a network visualization tool for users to draw their regulatory network by personal customization. Users can set the complexity of the network by limiting the number of nodes or edges. And the color of the nodes can be defined by different categories of the mRNAs and miRNAs, such as Gene-Ontology, pathway, and expression status. Users can also select to use network degree or network betweenness centrality to define the node size. And edges can be black or colored by the correlation. Purple edge means negative correlation (mostly found between miRNA and mRNA), and blue edge means positive correlation (found in PPI or miRNA-miRNA sponge effect). We can also add the protein-protein interactions (PPI) into the network. This result will show the cluster of genes regulated by some specific miRNAs. Additionally, miRNA-miRNA edges can be added by the "miRNA sponge" button, presenting some clusters of miRNAs that have the interactions via sponge effect.
miRNA-gene regulations
| Num | microRNA | Gene | miRNA log2FC | miRNA pvalue | Gene log2FC | Gene pvalue | Interaction | Correlation beta | Correlation P-value | PMID | Reported in cancer studies |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | hsa-miR-140-3p | ADAM17 | -0.3 | 0.12754 | 0.99 | 6.0E-5 | PITA; miRNATAP | -0.47 | 0 | NA | |
| 2 | hsa-miR-326 | ADAM17 | -0.43 | 0.26299 | 0.99 | 6.0E-5 | miRanda | -0.26 | 0 | 26111641 | Adam17 a Target of Mir 326 Promotes Emt Induced Cells Invasion in Lung Adenocarcinoma; RT-PCR and Western blot were used to detect the expression of miR-326 and ADAM17 in lung adenocarcinoma samples n=73; Furthermore we confirmed that ADAM17 is a target of miR-326 which is involved in EMT and cells invasion; These findings revealed that ADAM17 a target of miR-326 promoted EMT-induced cells invasion in lung adenocarcinoma |
| 3 | hsa-miR-338-3p | ADAM17 | -0.96 | 0.01915 | 0.99 | 6.0E-5 | PITA; miRanda; miRNATAP | -0.31 | 0 | 26617808 | MiR 338 3p inhibits the proliferation and migration of gastric cancer cells by targeting ADAM17; Moreover we identified A disintegrin and metalloproteinase 17 ADAM17 gene as potential target of miR-338-3p; Importantly ADAM17 rescued the miR-338-3p mediated inhibition of cell proliferation migration and invasion; Our study suggested that miR-338-3p is significantly decreased in gastric cancer and inhibits cell proliferation migration and invasion partially via the downregulation of ADAM17 |
| 4 | hsa-miR-7-1-3p | ADAM17 | 0.71 | 0.04123 | 0.99 | 6.0E-5 | mirMAP | -0.23 | 0 | NA | |
| 5 | hsa-miR-186-5p | CREBBP | 0.15 | 0.43471 | -0.11 | 0.45436 | mirMAP; miRNATAP | -0.14 | 0.00768 | NA | |
| 6 | hsa-miR-342-3p | CTBP2 | 0.32 | 0.26915 | -0.01 | 0.94438 | MirTarget; PITA; miRanda; miRNATAP | -0.29 | 0 | NA | |
| 7 | hsa-miR-107 | DLL1 | 0.9 | 5.0E-5 | 0.23 | 0.51166 | PITA; miRanda; miRNATAP | -0.3 | 0.00774 | NA | |
| 8 | hsa-miR-326 | DLL1 | -0.43 | 0.26299 | 0.23 | 0.51166 | miRanda | -0.21 | 0.00151 | NA | |
| 9 | hsa-miR-338-5p | DLL1 | -1.2 | 0.01003 | 0.23 | 0.51166 | PITA; miRNATAP | -0.22 | 2.0E-5 | NA | |
| 10 | hsa-miR-429 | DLL1 | 1.4 | 0.009 | 0.23 | 0.51166 | miRanda; miRNATAP | -0.12 | 0.00861 | NA | |
| 11 | hsa-let-7a-3p | DLL4 | 0.5 | 0.04111 | 0.32 | 0.35188 | miRNATAP | -0.33 | 0.001 | NA | |
| 12 | hsa-let-7b-3p | DLL4 | 0.22 | 0.29604 | 0.32 | 0.35188 | miRNATAP | -0.33 | 0.00644 | NA | |
| 13 | hsa-miR-125b-5p | DLL4 | -0.51 | 0.13327 | 0.32 | 0.35188 | miRNATAP | -0.29 | 4.0E-5 | NA | |
| 14 | hsa-miR-199a-3p | DLL4 | 0.85 | 0.00036 | 0.32 | 0.35188 | PITA | -0.31 | 0.00277 | NA | |
| 15 | hsa-miR-199b-3p | DLL4 | 0.86 | 0.00032 | 0.32 | 0.35188 | PITA | -0.31 | 0.00275 | NA | |
| 16 | hsa-miR-27a-3p | DLL4 | 1.3 | 0 | 0.32 | 0.35188 | miRNATAP | -0.5 | 0 | NA | |
| 17 | hsa-miR-27b-3p | DLL4 | 0.08 | 0.72527 | 0.32 | 0.35188 | miRNATAP | -0.57 | 0 | NA | |
| 18 | hsa-miR-944 | DLL4 | 3.33 | 0.01778 | 0.32 | 0.35188 | PITA; miRNATAP | -0.15 | 0 | NA | |
| 19 | hsa-miR-1254 | DTX1 | 0.94 | 0.06055 | -0.03 | 0.93372 | miRNATAP | -0.3 | 0 | NA | |
| 20 | hsa-miR-324-5p | DTX1 | 1.09 | 3.0E-5 | -0.03 | 0.93372 | PITA; miRanda | -0.45 | 1.0E-5 | NA | |
| 21 | hsa-miR-331-3p | DTX1 | 1.05 | 0.0001 | -0.03 | 0.93372 | PITA; miRNATAP | -0.41 | 3.0E-5 | NA | |
| 22 | hsa-miR-421 | DTX1 | 1.81 | 0 | -0.03 | 0.93372 | PITA; miRanda; miRNATAP | -0.25 | 0.00092 | NA | |
| 23 | hsa-miR-429 | DTX1 | 1.4 | 0.009 | -0.03 | 0.93372 | miRNATAP | -0.18 | 0.00019 | NA | |
| 24 | hsa-miR-484 | DTX1 | 0.71 | 0.00234 | -0.03 | 0.93372 | miRNATAP | -0.37 | 0.00134 | NA | |
| 25 | hsa-let-7e-5p | DTX2 | 0.41 | 0.19626 | 1.01 | 5.0E-5 | miRNATAP | -0.16 | 0.00444 | NA | |
| 26 | hsa-miR-107 | DTX3 | 0.9 | 5.0E-5 | -0.4 | 0.34209 | miRanda | -0.43 | 0.00139 | NA | |
| 27 | hsa-miR-148a-5p | DTX3 | -0.69 | 0.0793 | -0.4 | 0.34209 | mirMAP | -0.24 | 0.00121 | NA | |
| 28 | hsa-miR-3614-5p | DTX3 | 1.1 | 0.00706 | -0.4 | 0.34209 | MirTarget | -0.38 | 0 | NA | |
| 29 | hsa-let-7b-5p | DTX4 | 0.06 | 0.7814 | 0.44 | 0.32524 | miRNATAP | -0.67 | 1.0E-5 | NA | |
| 30 | hsa-miR-125b-5p | DTX4 | -0.51 | 0.13327 | 0.44 | 0.32524 | mirMAP; miRNATAP | -0.38 | 4.0E-5 | NA | |
| 31 | hsa-miR-136-5p | DTX4 | -0.39 | 0.19926 | 0.44 | 0.32524 | mirMAP | -0.3 | 0.00381 | NA | |
| 32 | hsa-miR-27a-3p | DTX4 | 1.3 | 0 | 0.44 | 0.32524 | miRNATAP | -0.77 | 0 | NA | |
| 33 | hsa-miR-27b-3p | DTX4 | 0.08 | 0.72527 | 0.44 | 0.32524 | miRNATAP | -0.76 | 0 | NA | |
| 34 | hsa-miR-365a-3p | DTX4 | -0.89 | 0.00255 | 0.44 | 0.32524 | mirMAP | -0.73 | 0 | NA | |
| 35 | hsa-miR-150-5p | DVL3 | 0.15 | 0.75372 | 0.51 | 0.01916 | MirTarget | -0.14 | 3.0E-5 | NA | |
| 36 | hsa-miR-29b-2-5p | DVL3 | -1.7 | 0 | 0.51 | 0.01916 | MirTarget | -0.26 | 0 | NA | |
| 37 | hsa-miR-30d-3p | DVL3 | -0.55 | 0.04337 | 0.51 | 0.01916 | MirTarget | -0.21 | 0.00028 | NA | |
| 38 | hsa-miR-338-3p | DVL3 | -0.96 | 0.01915 | 0.51 | 0.01916 | miRanda | -0.23 | 0 | NA | |
| 39 | hsa-miR-362-5p | DVL3 | -0.35 | 0.35491 | 0.51 | 0.01916 | PITA; TargetScan; miRNATAP | -0.21 | 0 | NA | |
| 40 | hsa-miR-106b-5p | EP300 | 1.71 | 0 | 0.03 | 0.83304 | miRNATAP | -0.1 | 0.00164 | NA | |
| 41 | hsa-miR-212-3p | EP300 | 0.96 | 0.00133 | 0.03 | 0.83304 | MirTarget; miRNATAP | -0.12 | 0.00033 | NA | |
| 42 | hsa-miR-26a-5p | EP300 | -0.38 | 0.04425 | 0.03 | 0.83304 | miRNATAP | -0.19 | 0.00038 | NA | |
| 43 | hsa-miR-26b-5p | EP300 | -0.3 | 0.16008 | 0.03 | 0.83304 | miRNAWalker2 validate; miRNATAP | -0.14 | 0.00299 | NA | |
| 44 | hsa-miR-342-3p | HDAC1 | 0.32 | 0.26915 | 0.78 | 0 | miRanda | -0.13 | 0.00172 | NA | |
| 45 | hsa-miR-145-5p | HDAC2 | -1.75 | 2.0E-5 | 0.54 | 0.00083 | miRNAWalker2 validate | -0.12 | 1.0E-5 | 23499894 | MiR 145 functions as a tumor suppressor by directly targeting histone deacetylase 2 in liver cancer; Ectopic expression of miRNA mimics evidenced that miR-145 suppresses HDAC2 expression in HCC cells; In conclusion we suggest that loss or suppression of miR-145 may cause aberrant overexpression of HDAC2 and promote HCC tumorigenesis |
| 46 | hsa-miR-195-3p | HDAC2 | -1.35 | 2.0E-5 | 0.54 | 0.00083 | mirMAP | -0.14 | 9.0E-5 | NA | |
| 47 | hsa-miR-30a-3p | HDAC2 | -1.73 | 0 | 0.54 | 0.00083 | mirMAP | -0.11 | 0.00143 | NA | |
| 48 | hsa-miR-30a-5p | HDAC2 | -1.72 | 0 | 0.54 | 0.00083 | mirMAP | -0.11 | 0.00069 | NA | |
| 49 | hsa-miR-129-5p | JAG1 | -2.57 | 0 | 0.96 | 0.00974 | PITA; miRanda; miRNATAP | -0.15 | 0.00169 | NA | |
| 50 | hsa-miR-140-3p | JAG1 | -0.3 | 0.12754 | 0.96 | 0.00974 | MirTarget; PITA; miRNATAP | -0.5 | 0.00033 | NA | |
| 51 | hsa-miR-186-5p | JAG1 | 0.15 | 0.43471 | 0.96 | 0.00974 | MirTarget; miRNATAP | -0.62 | 1.0E-5 | NA | |
| 52 | hsa-miR-200a-3p | JAG1 | 0.72 | 0.19391 | 0.96 | 0.00974 | miRNATAP | -0.14 | 0.0045 | NA | |
| 53 | hsa-miR-217 | JAG1 | -0.01 | 0.98082 | 0.96 | 0.00974 | PITA; miRanda | -0.16 | 0.00328 | NA | |
| 54 | hsa-miR-26a-5p | JAG1 | -0.38 | 0.04425 | 0.96 | 0.00974 | MirTarget; miRNATAP | -0.53 | 0.00017 | NA | |
| 55 | hsa-miR-26b-3p | JAG1 | -0.01 | 0.96577 | 0.96 | 0.00974 | miRNATAP | -0.3 | 0.00827 | NA | |
| 56 | hsa-miR-26b-5p | JAG1 | -0.3 | 0.16008 | 0.96 | 0.00974 | MirTarget; miRNATAP | -0.35 | 0.00639 | NA | |
| 57 | hsa-miR-335-3p | JAG1 | 2.52 | 0 | 0.96 | 0.00974 | MirTarget | -0.16 | 0.00723 | NA | |
| 58 | hsa-miR-335-5p | JAG1 | 1.6 | 6.0E-5 | 0.96 | 0.00974 | miRNATAP | -0.27 | 3.0E-5 | NA | |
| 59 | hsa-miR-361-3p | JAG1 | -0.13 | 0.56605 | 0.96 | 0.00974 | miRNATAP | -0.39 | 0.00176 | NA | |
| 60 | hsa-miR-375 | JAG1 | -2.95 | 0.0057 | 0.96 | 0.00974 | miRNAWalker2 validate | -0.16 | 0 | NA | |
| 61 | hsa-miR-429 | JAG1 | 1.4 | 0.009 | 0.96 | 0.00974 | miRanda | -0.13 | 0.00811 | NA | |
| 62 | hsa-miR-628-5p | JAG1 | -0.54 | 0.03779 | 0.96 | 0.00974 | MirTarget; PITA; miRNATAP | -0.41 | 6.0E-5 | NA | |
| 63 | hsa-miR-98-5p | JAG1 | 0.92 | 3.0E-5 | 0.96 | 0.00974 | miRNAWalker2 validate | -0.35 | 0.00367 | NA | |
| 64 | hsa-miR-140-3p | JAG2 | -0.3 | 0.12754 | 1.13 | 0.00219 | miRNATAP | -0.8 | 0 | NA | |
| 65 | hsa-miR-140-5p | JAG2 | 0.21 | 0.303 | 1.13 | 0.00219 | miRanda | -0.52 | 9.0E-5 | NA | |
| 66 | hsa-miR-30a-5p | JAG2 | -1.72 | 0 | 1.13 | 0.00219 | miRNATAP | -0.29 | 0.00011 | NA | |
| 67 | hsa-miR-30b-5p | JAG2 | -0.43 | 0.05936 | 1.13 | 0.00219 | miRNATAP | -0.39 | 0.00074 | NA | |
| 68 | hsa-miR-30c-5p | JAG2 | -0.98 | 5.0E-5 | 1.13 | 0.00219 | miRNATAP | -0.38 | 0.00042 | NA | |
| 69 | hsa-miR-30d-5p | JAG2 | -0.55 | 0.01401 | 1.13 | 0.00219 | miRNATAP | -0.51 | 1.0E-5 | NA | |
| 70 | hsa-miR-335-5p | JAG2 | 1.6 | 6.0E-5 | 1.13 | 0.00219 | miRNAWalker2 validate | -0.19 | 0.00353 | NA | |
| 71 | hsa-miR-501-3p | JAG2 | 0.73 | 0.02704 | 1.13 | 0.00219 | TargetScan; miRNATAP | -0.25 | 0.00174 | NA | |
| 72 | hsa-miR-502-3p | JAG2 | -0.16 | 0.55747 | 1.13 | 0.00219 | miRNATAP | -0.54 | 0 | NA | |
| 73 | hsa-miR-342-3p | KAT2A | 0.32 | 0.26915 | 0.95 | 0 | MirTarget; miRanda | -0.12 | 0.0077 | NA | |
| 74 | hsa-miR-106b-5p | KAT2B | 1.71 | 0 | -1.91 | 0 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.4 | 0 | NA | |
| 75 | hsa-miR-17-5p | KAT2B | 1.66 | 0 | -1.91 | 0 | MirTarget; TargetScan | -0.45 | 0 | 23095762 | miR 17 5p targets the p300/CBP associated factor and modulates androgen receptor transcriptional activity in cultured prostate cancer cells; Targeting of PCAF by miR-17-5p was evaluated using the luciferase reporter assay; Expression of PCAF in PCa cells was associated with the downregulation of miR-17-5p; Targeting of the 3'-untranslated region of PCAF mRNA by miR-17-5p caused translational suppression and RNA degradation and consequently modulation of AR transcriptional activity in PCa cells; PCAF is upregulated in cultured PCa cells and upregulation of PCAF is associated with the downregulation of miR-17-5p; Targeting of PCAF by miR-17-5p modulates AR transcriptional activity and cell growth in cultured PCa cells |
| 76 | hsa-miR-181a-5p | KAT2B | 1.39 | 0 | -1.91 | 0 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.32 | 0.00139 | NA | |
| 77 | hsa-miR-181b-5p | KAT2B | 1.64 | 0 | -1.91 | 0 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.3 | 0.00011 | NA | |
| 78 | hsa-miR-181d-5p | KAT2B | 0.65 | 0.01434 | -1.91 | 0 | MirTarget | -0.25 | 0.00538 | NA | |
| 79 | hsa-miR-19a-3p | KAT2B | 1.27 | 0.00011 | -1.91 | 0 | miRNAWalker2 validate | -0.27 | 0.00015 | NA | |
| 80 | hsa-miR-19b-3p | KAT2B | 0.76 | 0.00653 | -1.91 | 0 | miRNAWalker2 validate | -0.28 | 0.00111 | NA | |
| 81 | hsa-miR-20a-3p | KAT2B | 1.14 | 0.00045 | -1.91 | 0 | MirTarget | -0.42 | 0 | NA | |
| 82 | hsa-miR-20a-5p | KAT2B | 1.45 | 0 | -1.91 | 0 | MirTarget | -0.43 | 0 | NA | |
| 83 | hsa-miR-25-3p | KAT2B | 1.01 | 0.00042 | -1.91 | 0 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.3 | 0.00024 | NA | |
| 84 | hsa-miR-361-5p | KAT2B | 0.41 | 0.01813 | -1.91 | 0 | miRanda | -0.41 | 0.00393 | NA | |
| 85 | hsa-miR-590-3p | KAT2B | 1.12 | 0.00016 | -1.91 | 0 | MirTarget; miRanda; mirMAP; miRNATAP | -0.22 | 0.00595 | NA | |
| 86 | hsa-miR-590-5p | KAT2B | 1.04 | 0.00027 | -1.91 | 0 | miRanda | -0.33 | 6.0E-5 | NA | |
| 87 | hsa-miR-92a-3p | KAT2B | 1.22 | 1.0E-5 | -1.91 | 0 | miRNAWalker2 validate; MirTarget | -0.52 | 0 | NA | |
| 88 | hsa-miR-92b-3p | KAT2B | 1.34 | 0 | -1.91 | 0 | MirTarget | -0.49 | 0 | NA | |
| 89 | hsa-miR-93-5p | KAT2B | 1.75 | 0 | -1.91 | 0 | miRNAWalker2 validate; miRTarBase; MirTarget | -0.39 | 0 | NA | |
| 90 | hsa-miR-125a-5p | LFNG | -0.88 | 0.00021 | -0.13 | 0.76587 | MirTarget; PITA; miRanda; miRNATAP | -0.37 | 0.00675 | NA | |
| 91 | hsa-miR-125b-5p | LFNG | -0.51 | 0.13327 | -0.13 | 0.76587 | MirTarget; miRNATAP | -0.56 | 0 | NA | |
| 92 | hsa-miR-214-3p | LFNG | 1.01 | 0.00625 | -0.13 | 0.76587 | MirTarget | -0.23 | 0.00884 | NA | |
| 93 | hsa-miR-365a-3p | LFNG | -0.89 | 0.00255 | -0.13 | 0.76587 | mirMAP | -0.54 | 0 | NA | |
| 94 | hsa-miR-30b-5p | MAML1 | -0.43 | 0.05936 | 0.1 | 0.48957 | MirTarget | -0.13 | 0.00324 | NA | |
| 95 | hsa-miR-576-5p | MAML2 | 0.63 | 0.04171 | 0.01 | 0.97433 | mirMAP | -0.2 | 0.00345 | NA | |
| 96 | hsa-miR-205-5p | MFNG | 3.14 | 0.02932 | -0.44 | 0.22631 | MirTarget | -0.11 | 0 | NA | |
| 97 | hsa-miR-219a-5p | MFNG | 0.61 | 0.08545 | -0.44 | 0.22631 | MirTarget | -0.32 | 1.0E-5 | NA | |
| 98 | hsa-miR-2355-5p | MFNG | 0.59 | 0.12059 | -0.44 | 0.22631 | MirTarget | -0.22 | 0.00087 | NA | |
| 99 | hsa-miR-101-3p | NOTCH1 | -0.45 | 0.02834 | 1.03 | 0.00018 | MirTarget | -0.41 | 3.0E-5 | NA | |
| 100 | hsa-miR-139-5p | NOTCH1 | -1.83 | 0 | 1.03 | 0.00018 | PITA; miRanda; miRNATAP | -0.24 | 0.0001 | 27173050; 25149074; 26299922; 27738333; 24885920 | miR 139 5p sensitizes colorectal cancer cells to 5 fluorouracil by targeting NOTCH 1;MiR 139 5p inhibits migration and invasion of colorectal cancer by downregulating AMFR and NOTCH1; Mechanistic investigations revealed that miR-139-5p suppress CRC cell invasion and metastasis by targeting AMFR and NOTCH1;MiR 139 5p inhibits the biological function of breast cancer cells by targeting Notch1 and mediates chemosensitivity to docetaxel; MiR-139-5p expression in MCF-7 MCF-7/Doc cells and in selected breast cancer tissue samples was confirmed by real-time PCR; cell viability was analyzed by Cell Counting Kit-8 assay; apoptosis and cell cycle were analyzed by flow cytometry; control of metastasis and invasion of breast cancer cells was measured by transwell assay; expression of Notch1 was measured by western blot; a luciferase reporter vector was constructed to identify the miR-139-5p target gene; MiR-139-5p not only attenuated the development of breast cancer cells but also mediated drug-resistance by regulating the expression of the downstream target gene Notch1;MiR 139 5p reverses CD44+/CD133+ associated multidrug resistance by downregulating NOTCH1 in colorectal carcinoma cells; We also identified NOTCH1 an important protein for stem cell maintenance and function as a direct target of miR-139-5p both in vitro and in a knockout mouse model; Notch1 expression was upregulated in tumor samples and inversely correlated with expression of miR-139-5p; Silencing NOTCH1 exerted an effect similar to overexpression of miR-139-5p by inhibiting the CD44+ and CD133+ population and reversing the drug-resistant phenotype; In conclusion miR-139-5p downregulated NOTCH1 signaling to reverse CD44+/CD133+-associated MDR in colorectal cancer cells;microRNA 139 5p exerts tumor suppressor function by targeting NOTCH1 in colorectal cancer |
| 101 | hsa-miR-30a-5p | NOTCH1 | -1.72 | 0 | 1.03 | 0.00018 | miRNAWalker2 validate; miRTarBase; miRNATAP | -0.24 | 3.0E-5 | NA | |
| 102 | hsa-miR-30b-5p | NOTCH1 | -0.43 | 0.05936 | 1.03 | 0.00018 | miRNATAP | -0.36 | 4.0E-5 | NA | |
| 103 | hsa-miR-30c-5p | NOTCH1 | -0.98 | 5.0E-5 | 1.03 | 0.00018 | miRNATAP | -0.33 | 6.0E-5 | 23974200 | We identified NOTCH1 as a direct target of miR-30c; Finally a block of miR-30c prevents C/EBPα-induced downregulation of Notch1 protein and leads to a reduced CD11b expression in myeloid differentiation; Our study presents the first evidence that C/EBPα miR-30c and Notch1 together play a critical role in granulocytic differentiation and AML and particularly in AML with CEBPA mutations |
| 104 | hsa-miR-30d-5p | NOTCH1 | -0.55 | 0.01401 | 1.03 | 0.00018 | miRNATAP | -0.28 | 0.00181 | NA | |
| 105 | hsa-miR-32-5p | NOTCH1 | 0.42 | 0.10646 | 1.03 | 0.00018 | miRNATAP | -0.25 | 0.00133 | NA | |
| 106 | hsa-miR-326 | NOTCH1 | -0.43 | 0.26299 | 1.03 | 0.00018 | miRNAWalker2 validate; miRTarBase | -0.17 | 0.00149 | NA | |
| 107 | hsa-miR-103a-3p | NOTCH2 | 0.84 | 0 | -0.26 | 0.28775 | MirTarget | -0.37 | 0.00028 | NA | |
| 108 | hsa-miR-106a-5p | NOTCH2 | -0.17 | 0.64287 | -0.26 | 0.28775 | miRNAWalker2 validate | -0.13 | 0.00358 | NA | |
| 109 | hsa-miR-107 | NOTCH2 | 0.9 | 5.0E-5 | -0.26 | 0.28775 | miRNAWalker2 validate; MirTarget; miRanda; miRNATAP | -0.36 | 0 | NA | |
| 110 | hsa-miR-130b-5p | NOTCH2 | 0.7 | 0.05101 | -0.26 | 0.28775 | miRNAWalker2 validate; MirTarget | -0.18 | 0.00012 | NA | |
| 111 | hsa-miR-15a-5p | NOTCH2 | 1.04 | 0 | -0.26 | 0.28775 | MirTarget | -0.38 | 0 | NA | |
| 112 | hsa-miR-15b-5p | NOTCH2 | 1.62 | 0 | -0.26 | 0.28775 | MirTarget | -0.22 | 0.00025 | NA | |
| 113 | hsa-miR-16-1-3p | NOTCH2 | 1.43 | 0 | -0.26 | 0.28775 | mirMAP | -0.19 | 0.00335 | NA | |
| 114 | hsa-miR-16-5p | NOTCH2 | 1.01 | 1.0E-5 | -0.26 | 0.28775 | miRNAWalker2 validate; MirTarget | -0.39 | 0 | NA | |
| 115 | hsa-miR-17-5p | NOTCH2 | 1.66 | 0 | -0.26 | 0.28775 | miRNAWalker2 validate | -0.24 | 1.0E-5 | NA | |
| 116 | hsa-miR-183-5p | NOTCH2 | 1.66 | 0.00052 | -0.26 | 0.28775 | miRNAWalker2 validate | -0.12 | 0.00103 | NA | |
| 117 | hsa-miR-185-5p | NOTCH2 | 1.14 | 0 | -0.26 | 0.28775 | MirTarget | -0.28 | 0.00099 | NA | |
| 118 | hsa-miR-186-5p | NOTCH2 | 0.15 | 0.43471 | -0.26 | 0.28775 | mirMAP | -0.32 | 0.0006 | NA | |
| 119 | hsa-miR-18a-3p | NOTCH2 | 2.35 | 0 | -0.26 | 0.28775 | MirTarget | -0.16 | 4.0E-5 | NA | |
| 120 | hsa-miR-200a-5p | NOTCH2 | 1.5 | 0.00264 | -0.26 | 0.28775 | mirMAP | -0.13 | 9.0E-5 | NA | |
| 121 | hsa-miR-200b-5p | NOTCH2 | 0.97 | 0.05305 | -0.26 | 0.28775 | mirMAP | -0.16 | 1.0E-5 | NA | |
| 122 | hsa-miR-25-3p | NOTCH2 | 1.01 | 0.00042 | -0.26 | 0.28775 | miRNAWalker2 validate | -0.19 | 0.00136 | NA | |
| 123 | hsa-miR-29b-3p | NOTCH2 | -0.23 | 0.36746 | -0.26 | 0.28775 | MirTarget | -0.19 | 0.00526 | NA | |
| 124 | hsa-miR-30b-5p | NOTCH2 | -0.43 | 0.05936 | -0.26 | 0.28775 | mirMAP | -0.24 | 0.00141 | NA | |
| 125 | hsa-miR-34a-5p | NOTCH2 | 0.79 | 0.00024 | -0.26 | 0.28775 | miRNAWalker2 validate; miRNATAP | -0.22 | 0.00531 | NA | |
| 126 | hsa-miR-423-5p | NOTCH2 | 0.83 | 0.00079 | -0.26 | 0.28775 | MirTarget | -0.25 | 0.00035 | NA | |
| 127 | hsa-miR-424-5p | NOTCH2 | 1.09 | 0.00042 | -0.26 | 0.28775 | MirTarget | -0.22 | 7.0E-5 | NA | |
| 128 | hsa-miR-429 | NOTCH2 | 1.4 | 0.009 | -0.26 | 0.28775 | miRNATAP | -0.15 | 0 | NA | |
| 129 | hsa-miR-577 | NOTCH2 | 0.91 | 0.22561 | -0.26 | 0.28775 | mirMAP | -0.13 | 0 | NA | |
| 130 | hsa-miR-590-3p | NOTCH2 | 1.12 | 0.00016 | -0.26 | 0.28775 | miRanda | -0.2 | 0.00043 | NA | |
| 131 | hsa-miR-590-5p | NOTCH2 | 1.04 | 0.00027 | -0.26 | 0.28775 | miRanda | -0.21 | 0.00041 | NA | |
| 132 | hsa-miR-7-1-3p | NOTCH2 | 0.71 | 0.04123 | -0.26 | 0.28775 | MirTarget; mirMAP | -0.21 | 2.0E-5 | NA | |
| 133 | hsa-miR-92a-3p | NOTCH2 | 1.22 | 1.0E-5 | -0.26 | 0.28775 | miRNAWalker2 validate | -0.29 | 0 | NA | |
| 134 | hsa-miR-940 | NOTCH2 | 1.64 | 0.00069 | -0.26 | 0.28775 | MirTarget | -0.13 | 0.00177 | NA | |
| 135 | hsa-miR-150-5p | NOTCH3 | 0.15 | 0.75372 | 1.32 | 0.00125 | miRNAWalker2 validate; miRTarBase | -0.16 | 0.00986 | NA | |
| 136 | hsa-miR-200a-3p | NOTCH3 | 0.72 | 0.19391 | 1.32 | 0.00125 | mirMAP | -0.29 | 0 | NA | |
| 137 | hsa-miR-335-5p | NOTCH3 | 1.6 | 6.0E-5 | 1.32 | 0.00125 | miRNAWalker2 validate | -0.27 | 0.00023 | NA | |
| 138 | hsa-miR-491-5p | NOTCH3 | -0.69 | 0.03136 | 1.32 | 0.00125 | MirTarget; miRanda | -0.3 | 0.00117 | NA | |
| 139 | hsa-miR-660-5p | NOTCH3 | -0.02 | 0.93887 | 1.32 | 0.00125 | MirTarget | -0.76 | 0 | NA | |
| 140 | hsa-miR-7-5p | NOTCH3 | 1.64 | 0.01244 | 1.32 | 0.00125 | MirTarget | -0.29 | 0 | NA | |
| 141 | hsa-miR-324-5p | NOTCH4 | 1.09 | 3.0E-5 | -0.76 | 0.0205 | miRanda | -0.28 | 0.00164 | NA | |
| 142 | hsa-miR-34c-5p | NOTCH4 | 2.21 | 0.00038 | -0.76 | 0.0205 | miRNAWalker2 validate; miRTarBase; miRanda; miRNATAP | -0.17 | 0 | 22074923 | Ectopic expression of miR-34c reduced the self-renewal of BT-ICs inhibited epithelial-mesenchymal transition and suppressed migration of the tumor cells via silencing target gene Notch4 |
| 143 | hsa-miR-103a-3p | NUMB | 0.84 | 0 | -0.06 | 0.68101 | miRNAWalker2 validate | -0.17 | 0.00578 | NA | |
| 144 | hsa-miR-374b-5p | NUMB | -0.19 | 0.3948 | -0.06 | 0.68101 | miRNATAP | -0.15 | 0.00115 | NA | |
| 145 | hsa-let-7g-5p | NUMBL | 0.33 | 0.24114 | 0.2 | 0.4531 | miRNATAP | -0.29 | 1.0E-5 | NA | |
| 146 | hsa-miR-140-5p | NUMBL | 0.21 | 0.303 | 0.2 | 0.4531 | MirTarget; miRanda; miRNATAP | -0.41 | 1.0E-5 | NA | |
| 147 | hsa-miR-141-3p | NUMBL | 1.46 | 0.00116 | 0.2 | 0.4531 | TargetScan | -0.13 | 0.00179 | NA | |
| 148 | hsa-miR-142-3p | NUMBL | 1.32 | 0.00015 | 0.2 | 0.4531 | miRNATAP | -0.19 | 0.00022 | NA | |
| 149 | hsa-miR-194-3p | NUMBL | 0.26 | 0.82181 | 0.2 | 0.4531 | MirTarget | -0.13 | 0 | NA | |
| 150 | hsa-miR-338-3p | NUMBL | -0.96 | 0.01915 | 0.2 | 0.4531 | miRanda | -0.27 | 0 | NA |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
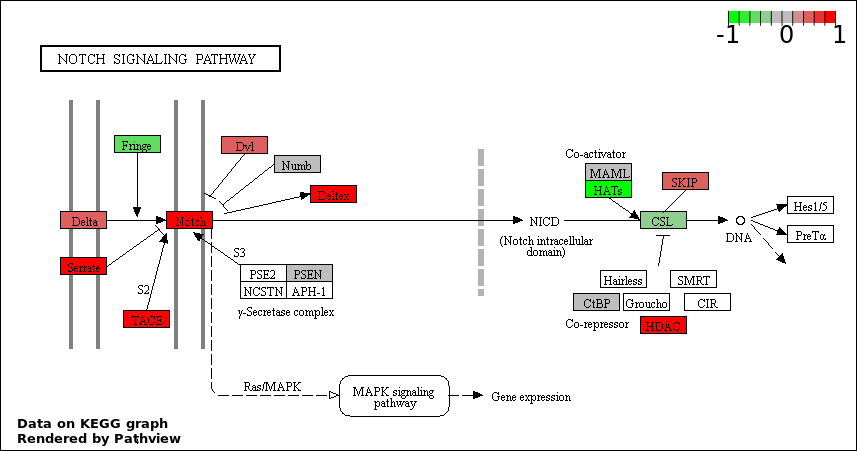
| 1 | NOTCH SIGNALING PATHWAY | 22 | 114 | 9.556e-44 | 4.446e-40 |
| 2 | NOTCH RECEPTOR PROCESSING | 9 | 16 | 1.626e-22 | 3.783e-19 |
| 3 | REGULATION OF NOTCH SIGNALING PATHWAY | 9 | 67 | 5.778e-16 | 8.962e-13 |
| 4 | TRANSCRIPTION INITIATION FROM RNA POLYMERASE II PROMOTER | 10 | 153 | 1.97e-14 | 2.292e-11 |
| 5 | POSITIVE REGULATION OF NOTCH SIGNALING PATHWAY | 7 | 34 | 5.42e-14 | 5.044e-11 |
| 6 | EPITHELIUM DEVELOPMENT | 16 | 945 | 8.4e-14 | 6.514e-11 |
| 7 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 16 | 1004 | 2.135e-13 | 1.419e-10 |
| 8 | POSITIVE REGULATION OF GENE EXPRESSION | 19 | 1733 | 3.057e-13 | 1.684e-10 |
| 9 | DNA TEMPLATED TRANSCRIPTION INITIATION | 10 | 202 | 3.257e-13 | 1.684e-10 |
| 10 | TISSUE DEVELOPMENT | 18 | 1518 | 5.036e-13 | 2.343e-10 |
| 11 | CELL FATE COMMITMENT | 10 | 227 | 1.047e-12 | 4.431e-10 |
| 12 | REGULATION OF NERVOUS SYSTEM DEVELOPMENT | 14 | 750 | 1.416e-12 | 5.489e-10 |
| 13 | ANATOMICAL STRUCTURE FORMATION INVOLVED IN MORPHOGENESIS | 15 | 957 | 2.074e-12 | 7.423e-10 |
| 14 | REGULATION OF CELL DIFFERENTIATION | 17 | 1492 | 6.089e-12 | 2.024e-09 |
| 15 | REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 18 | 1784 | 7.833e-12 | 2.43e-09 |
| 16 | POSITIVE REGULATION OF BIOSYNTHETIC PROCESS | 18 | 1805 | 9.543e-12 | 2.775e-09 |
| 17 | MORPHOGENESIS OF AN EPITHELIUM | 11 | 400 | 1.056e-11 | 2.891e-09 |
| 18 | LEUKOCYTE DIFFERENTIATION | 10 | 292 | 1.277e-11 | 3.3e-09 |
| 19 | EMBRYO DEVELOPMENT | 14 | 894 | 1.5e-11 | 3.672e-09 |
| 20 | TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 13 | 724 | 1.862e-11 | 4.332e-09 |
| 21 | LYMPHOCYTE DIFFERENTIATION | 9 | 209 | 2.066e-11 | 4.577e-09 |
| 22 | IMMUNE SYSTEM DEVELOPMENT | 12 | 582 | 2.807e-11 | 5.679e-09 |
| 23 | CELL SURFACE RECEPTOR SIGNALING PATHWAY INVOLVED IN HEART DEVELOPMENT | 5 | 16 | 2.752e-11 | 5.679e-09 |
| 24 | REGULATION OF MULTICELLULAR ORGANISMAL DEVELOPMENT | 17 | 1672 | 3.751e-11 | 7.272e-09 |
| 25 | HEART DEVELOPMENT | 11 | 466 | 5.435e-11 | 9.367e-09 |
| 26 | CARDIOVASCULAR SYSTEM DEVELOPMENT | 13 | 788 | 5.346e-11 | 9.367e-09 |
| 27 | CIRCULATORY SYSTEM DEVELOPMENT | 13 | 788 | 5.346e-11 | 9.367e-09 |
| 28 | LYMPHOCYTE ACTIVATION | 10 | 342 | 6.047e-11 | 1.005e-08 |
| 29 | B CELL DIFFERENTIATION | 7 | 89 | 6.551e-11 | 1.051e-08 |
| 30 | REGULATION OF CELL DEVELOPMENT | 13 | 836 | 1.113e-10 | 1.726e-08 |
| 31 | ORGAN MORPHOGENESIS | 13 | 841 | 1.198e-10 | 1.798e-08 |
| 32 | POSITIVE REGULATION OF CELL COMMUNICATION | 16 | 1532 | 1.295e-10 | 1.883e-08 |
| 33 | TISSUE MORPHOGENESIS | 11 | 533 | 2.271e-10 | 3.203e-08 |
| 34 | TUBE DEVELOPMENT | 11 | 552 | 3.292e-10 | 4.505e-08 |
| 35 | POSITIVE REGULATION OF RESPONSE TO STIMULUS | 17 | 1929 | 3.582e-10 | 4.762e-08 |
| 36 | LEUKOCYTE ACTIVATION | 10 | 414 | 3.9e-10 | 5.041e-08 |
| 37 | PATTERN SPECIFICATION PROCESS | 10 | 418 | 4.282e-10 | 5.385e-08 |
| 38 | NEUROGENESIS | 15 | 1402 | 4.642e-10 | 5.684e-08 |
| 39 | SOMITOGENESIS | 6 | 62 | 4.799e-10 | 5.725e-08 |
| 40 | NEURON FATE COMMITMENT | 6 | 67 | 7.748e-10 | 9.013e-08 |
| 41 | HEART MORPHOGENESIS | 8 | 212 | 8.935e-10 | 1.014e-07 |
| 42 | B CELL ACTIVATION | 7 | 132 | 1.071e-09 | 1.186e-07 |
| 43 | NEGATIVE REGULATION OF DEVELOPMENTAL PROCESS | 12 | 801 | 1.098e-09 | 1.188e-07 |
| 44 | CARDIAC CHAMBER DEVELOPMENT | 7 | 144 | 1.971e-09 | 2.038e-07 |
| 45 | SOMITE DEVELOPMENT | 6 | 78 | 1.971e-09 | 2.038e-07 |
| 46 | MATURE B CELL DIFFERENTIATION INVOLVED IN IMMUNE RESPONSE | 4 | 13 | 3.343e-09 | 3.381e-07 |
| 47 | SEGMENTATION | 6 | 89 | 4.406e-09 | 4.362e-07 |
| 48 | ENDOTHELIUM DEVELOPMENT | 6 | 90 | 4.716e-09 | 4.572e-07 |
| 49 | EMBRYONIC MORPHOGENESIS | 10 | 539 | 4.958e-09 | 4.708e-07 |
| 50 | REGULATION OF NEURON DIFFERENTIATION | 10 | 554 | 6.444e-09 | 5.879e-07 |
| 51 | EPITHELIAL CELL FATE COMMITMENT | 4 | 15 | 6.368e-09 | 5.879e-07 |
| 52 | NEGATIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER | 11 | 740 | 7.111e-09 | 6.363e-07 |
| 53 | CELL ACTIVATION | 10 | 568 | 8.175e-09 | 7.177e-07 |
| 54 | REGULATION OF BINDING | 8 | 283 | 8.661e-09 | 7.463e-07 |
| 55 | MATURE B CELL DIFFERENTIATION | 4 | 17 | 1.108e-08 | 9.373e-07 |
| 56 | CARDIAC CHAMBER MORPHOGENESIS | 6 | 104 | 1.132e-08 | 9.408e-07 |
| 57 | NEGATIVE REGULATION OF NEURON DIFFERENTIATION | 7 | 191 | 1.406e-08 | 1.143e-06 |
| 58 | ARTERY MORPHOGENESIS | 5 | 51 | 1.425e-08 | 1.143e-06 |
| 59 | ANTERIOR POSTERIOR PATTERN SPECIFICATION | 7 | 194 | 1.565e-08 | 1.229e-06 |
| 60 | NEGATIVE REGULATION OF CELL DIFFERENTIATION | 10 | 609 | 1.585e-08 | 1.229e-06 |
| 61 | REGIONALIZATION | 8 | 311 | 1.806e-08 | 1.378e-06 |
| 62 | POSITIVE REGULATION OF CELL DIFFERENTIATION | 11 | 823 | 2.133e-08 | 1.601e-06 |
| 63 | TUBE MORPHOGENESIS | 8 | 323 | 2.423e-08 | 1.79e-06 |
| 64 | SKIN DEVELOPMENT | 7 | 211 | 2.793e-08 | 2.031e-06 |
| 65 | POSITIVE REGULATION OF BINDING | 6 | 127 | 3.763e-08 | 2.694e-06 |
| 66 | SENSORY ORGAN DEVELOPMENT | 9 | 493 | 3.887e-08 | 2.741e-06 |
| 67 | NEURON DIFFERENTIATION | 11 | 874 | 3.953e-08 | 2.745e-06 |
| 68 | EPITHELIAL CELL DIFFERENTIATION | 9 | 495 | 4.025e-08 | 2.754e-06 |
| 69 | POSITIVE REGULATION OF MULTICELLULAR ORGANISMAL PROCESS | 13 | 1395 | 5.502e-08 | 3.71e-06 |
| 70 | POSITIVE REGULATION OF DEVELOPMENTAL PROCESS | 12 | 1142 | 5.813e-08 | 3.864e-06 |
| 71 | EPIDERMAL CELL DIFFERENTIATION | 6 | 142 | 7.328e-08 | 4.803e-06 |
| 72 | SKIN EPIDERMIS DEVELOPMENT | 5 | 71 | 7.727e-08 | 4.993e-06 |
| 73 | ENDOTHELIAL CELL DIFFERENTIATION | 5 | 72 | 8.294e-08 | 5.287e-06 |
| 74 | AUDITORY RECEPTOR CELL DIFFERENTIATION | 4 | 28 | 9.419e-08 | 5.923e-06 |
| 75 | EPIDERMIS DEVELOPMENT | 7 | 253 | 9.682e-08 | 6.007e-06 |
| 76 | ARTERY DEVELOPMENT | 5 | 75 | 1.02e-07 | 6.244e-06 |
| 77 | EMBRYO DEVELOPMENT ENDING IN BIRTH OR EGG HATCHING | 9 | 554 | 1.053e-07 | 6.363e-06 |
| 78 | REGULATION OF MUSCLE CELL DIFFERENTIATION | 6 | 152 | 1.098e-07 | 6.553e-06 |
| 79 | NEGATIVE REGULATION OF NERVOUS SYSTEM DEVELOPMENT | 7 | 262 | 1.228e-07 | 7.235e-06 |
| 80 | REGULATION OF CELL PROLIFERATION | 13 | 1496 | 1.253e-07 | 7.285e-06 |
| 81 | REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER IN RESPONSE TO HYPOXIA | 4 | 32 | 1.647e-07 | 9.462e-06 |
| 82 | MUSCLE TISSUE DEVELOPMENT | 7 | 275 | 1.707e-07 | 9.687e-06 |
| 83 | CARDIAC SEPTUM DEVELOPMENT | 5 | 85 | 1.917e-07 | 1.05e-05 |
| 84 | REGULATION OF STRIATED MUSCLE CELL DIFFERENTIATION | 5 | 85 | 1.917e-07 | 1.05e-05 |
| 85 | REGULATION OF ANATOMICAL STRUCTURE MORPHOGENESIS | 11 | 1021 | 1.917e-07 | 1.05e-05 |
| 86 | MUSCLE STRUCTURE DEVELOPMENT | 8 | 432 | 2.266e-07 | 1.226e-05 |
| 87 | HAIR CELL DIFFERENTIATION | 4 | 35 | 2.391e-07 | 1.264e-05 |
| 88 | B CELL ACTIVATION INVOLVED IN IMMUNE RESPONSE | 4 | 35 | 2.391e-07 | 1.264e-05 |
| 89 | POSITIVE REGULATION OF NERVOUS SYSTEM DEVELOPMENT | 8 | 437 | 2.473e-07 | 1.293e-05 |
| 90 | ANGIOGENESIS | 7 | 293 | 2.623e-07 | 1.356e-05 |
| 91 | MYOBLAST DIFFERENTIATION | 4 | 37 | 3.009e-07 | 1.538e-05 |
| 92 | NEGATIVE REGULATION OF CELL DEVELOPMENT | 7 | 303 | 3.291e-07 | 1.664e-05 |
| 93 | CARDIOCYTE DIFFERENTIATION | 5 | 96 | 3.53e-07 | 1.766e-05 |
| 94 | RESPONSE TO OXYGEN LEVELS | 7 | 311 | 3.923e-07 | 1.921e-05 |
| 95 | LYMPHOCYTE ACTIVATION INVOLVED IN IMMUNE RESPONSE | 5 | 98 | 3.914e-07 | 1.921e-05 |
| 96 | VASCULATURE DEVELOPMENT | 8 | 469 | 4.23e-07 | 2.05e-05 |
| 97 | IMMUNE SYSTEM PROCESS | 14 | 1984 | 4.434e-07 | 2.108e-05 |
| 98 | POSITIVE REGULATION OF CELL DEVELOPMENT | 8 | 472 | 4.439e-07 | 2.108e-05 |
| 99 | EAR DEVELOPMENT | 6 | 195 | 4.781e-07 | 2.247e-05 |
| 100 | MORPHOGENESIS OF AN EPITHELIAL SHEET | 4 | 43 | 5.586e-07 | 2.548e-05 |
| 101 | CELL FATE DETERMINATION | 4 | 43 | 5.586e-07 | 2.548e-05 |
| 102 | VENTRICULAR TRABECULA MYOCARDIUM MORPHOGENESIS | 3 | 11 | 5.517e-07 | 2.548e-05 |
| 103 | CELL DEVELOPMENT | 12 | 1426 | 6.497e-07 | 2.935e-05 |
| 104 | REGULATION OF TIMING OF CELL DIFFERENTIATION | 3 | 12 | 7.348e-07 | 3.226e-05 |
| 105 | COLUMNAR CUBOIDAL EPITHELIAL CELL DIFFERENTIATION | 5 | 111 | 7.283e-07 | 3.226e-05 |
| 106 | REGULATION OF DEVELOPMENT HETEROCHRONIC | 3 | 12 | 7.348e-07 | 3.226e-05 |
| 107 | CARDIAC SEPTUM MORPHOGENESIS | 4 | 49 | 9.528e-07 | 4.143e-05 |
| 108 | FOREBRAIN DEVELOPMENT | 7 | 357 | 9.899e-07 | 4.265e-05 |
| 109 | BLOOD VESSEL MORPHOGENESIS | 7 | 364 | 1.127e-06 | 4.767e-05 |
| 110 | MECHANORECEPTOR DIFFERENTIATION | 4 | 51 | 1.121e-06 | 4.767e-05 |
| 111 | NEGATIVE REGULATION OF MULTICELLULAR ORGANISMAL PROCESS | 10 | 983 | 1.355e-06 | 5.68e-05 |
| 112 | N TERMINAL PROTEIN AMINO ACID ACETYLATION | 3 | 15 | 1.515e-06 | 6.294e-05 |
| 113 | MAINTENANCE OF CELL NUMBER | 5 | 132 | 1.718e-06 | 7.074e-05 |
| 114 | MORPHOGENESIS OF AN ENDOTHELIUM | 3 | 16 | 1.863e-06 | 7.602e-05 |
| 115 | CELL ACTIVATION INVOLVED IN IMMUNE RESPONSE | 5 | 139 | 2.216e-06 | 8.967e-05 |
| 116 | EMBRYONIC DIGIT MORPHOGENESIS | 4 | 61 | 2.317e-06 | 9.135e-05 |
| 117 | CARDIAC MUSCLE TISSUE DEVELOPMENT | 5 | 140 | 2.296e-06 | 9.135e-05 |
| 118 | REGULATION OF VIRAL TRANSCRIPTION | 4 | 61 | 2.317e-06 | 9.135e-05 |
| 119 | CARDIAC VENTRICLE MORPHOGENESIS | 4 | 62 | 2.474e-06 | 9.672e-05 |
| 120 | NEUROEPITHELIAL CELL DIFFERENTIATION | 4 | 63 | 2.639e-06 | 0.0001023 |
| 121 | REGULATION OF PROTEIN ACETYLATION | 4 | 64 | 2.811e-06 | 0.0001081 |
| 122 | NEURONAL STEM CELL POPULATION MAINTENANCE | 3 | 19 | 3.213e-06 | 0.0001225 |
| 123 | REGULATION OF DNA TEMPLATED TRANSCRIPTION IN RESPONSE TO STRESS | 4 | 67 | 3.381e-06 | 0.0001279 |
| 124 | CELL FATE SPECIFICATION | 4 | 71 | 4.267e-06 | 0.0001601 |
| 125 | CENTRAL NERVOUS SYSTEM DEVELOPMENT | 9 | 872 | 4.63e-06 | 0.0001723 |
| 126 | MODULATION OF TRANSCRIPTION IN OTHER ORGANISM INVOLVED IN SYMBIOTIC INTERACTION | 3 | 22 | 5.09e-06 | 0.0001836 |
| 127 | MEMBRANE PROTEIN ECTODOMAIN PROTEOLYSIS | 3 | 22 | 5.09e-06 | 0.0001836 |
| 128 | SOMATIC STEM CELL DIVISION | 3 | 22 | 5.09e-06 | 0.0001836 |
| 129 | AORTA MORPHOGENESIS | 3 | 22 | 5.09e-06 | 0.0001836 |
| 130 | RHYTHMIC PROCESS | 6 | 298 | 5.599e-06 | 0.0002004 |
| 131 | APPENDAGE DEVELOPMENT | 5 | 169 | 5.774e-06 | 0.0002035 |
| 132 | LIMB DEVELOPMENT | 5 | 169 | 5.774e-06 | 0.0002035 |
| 133 | IMMUNE EFFECTOR PROCESS | 7 | 486 | 7.598e-06 | 0.0002658 |
| 134 | HAIR CYCLE | 4 | 83 | 7.966e-06 | 0.0002746 |
| 135 | MOLTING CYCLE | 4 | 83 | 7.966e-06 | 0.0002746 |
| 136 | NEGATIVE REGULATION OF GENE EXPRESSION | 11 | 1493 | 8.089e-06 | 0.0002767 |
| 137 | HEART TRABECULA MORPHOGENESIS | 3 | 26 | 8.557e-06 | 0.0002885 |
| 138 | N TERMINAL PROTEIN AMINO ACID MODIFICATION | 3 | 26 | 8.557e-06 | 0.0002885 |
| 139 | POSITIVE REGULATION OF CHROMATIN MODIFICATION | 4 | 85 | 8.758e-06 | 0.0002932 |
| 140 | HEAD DEVELOPMENT | 8 | 709 | 9.17e-06 | 0.0003048 |
| 141 | NEGATIVE REGULATION OF NITROGEN COMPOUND METABOLIC PROCESS | 11 | 1517 | 9.422e-06 | 0.0003109 |
| 142 | TISSUE REMODELING | 4 | 87 | 9.607e-06 | 0.0003129 |
| 143 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER INVOLVED IN CELLULAR RESPONSE TO CHEMICAL STIMULUS | 3 | 27 | 9.617e-06 | 0.0003129 |
| 144 | NEGATIVE REGULATION OF NOTCH SIGNALING PATHWAY | 3 | 28 | 1.076e-05 | 0.0003453 |
| 145 | REGULATION OF SPROUTING ANGIOGENESIS | 3 | 28 | 1.076e-05 | 0.0003453 |
| 146 | STEM CELL DIVISION | 3 | 29 | 1.199e-05 | 0.0003821 |
| 147 | COVALENT CHROMATIN MODIFICATION | 6 | 345 | 1.29e-05 | 0.0004084 |
| 148 | CARDIAC ATRIUM DEVELOPMENT | 3 | 31 | 1.472e-05 | 0.0004627 |
| 149 | ENDOCARDIAL CUSHION DEVELOPMENT | 3 | 32 | 1.622e-05 | 0.0005032 |
| 150 | BLOOD VESSEL REMODELING | 3 | 32 | 1.622e-05 | 0.0005032 |
| 151 | POSITIVE REGULATION OF TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 4 | 100 | 1.669e-05 | 0.0005143 |
| 152 | HEART VALVE DEVELOPMENT | 3 | 34 | 1.953e-05 | 0.0005979 |
| 153 | CARDIAC VENTRICLE DEVELOPMENT | 4 | 106 | 2.101e-05 | 0.0006388 |
| 154 | MEMBRANE PROTEIN PROTEOLYSIS | 3 | 35 | 2.134e-05 | 0.0006447 |
| 155 | POSITIVE REGULATION OF PROTEIN ACETYLATION | 3 | 36 | 2.325e-05 | 0.0006981 |
| 156 | POSITIVE REGULATION OF CELL PROLIFERATION | 8 | 814 | 2.493e-05 | 0.0007435 |
| 157 | REGULATION OF CELLULAR RESPONSE TO GROWTH FACTOR STIMULUS | 5 | 229 | 2.511e-05 | 0.0007441 |
| 158 | NEGATIVE REGULATION OF EPITHELIAL CELL DIFFERENTIATION | 3 | 37 | 2.528e-05 | 0.0007445 |
| 159 | REGULATION OF EMBRYONIC DEVELOPMENT | 4 | 114 | 2.797e-05 | 0.0008186 |
| 160 | NEPHRON DEVELOPMENT | 4 | 115 | 2.895e-05 | 0.0008419 |
| 161 | POSITIVE REGULATION OF VIRAL TRANSCRIPTION | 3 | 39 | 2.967e-05 | 0.000847 |
| 162 | TRABECULA MORPHOGENESIS | 3 | 39 | 2.967e-05 | 0.000847 |
| 163 | MUSCLE CELL DIFFERENTIATION | 5 | 237 | 2.959e-05 | 0.000847 |
| 164 | IMMUNE RESPONSE | 9 | 1100 | 2.99e-05 | 0.0008484 |
| 165 | POSITIVE REGULATION OF GROWTH | 5 | 238 | 3.02e-05 | 0.0008515 |
| 166 | SENSORY ORGAN MORPHOGENESIS | 5 | 239 | 3.081e-05 | 0.0008636 |
| 167 | AORTA DEVELOPMENT | 3 | 41 | 3.454e-05 | 0.0009623 |
| 168 | REGULATION OF EPITHELIAL CELL DIFFERENTIATION | 4 | 122 | 3.65e-05 | 0.001011 |
| 169 | T CELL DIFFERENTIATION | 4 | 123 | 3.769e-05 | 0.001038 |
| 170 | BETA CATENIN TCF COMPLEX ASSEMBLY | 3 | 43 | 3.99e-05 | 0.001092 |
| 171 | NEGATIVE REGULATION OF CELL DEATH | 8 | 872 | 4.077e-05 | 0.001109 |
| 172 | LEUKOCYTE CELL CELL ADHESION | 5 | 255 | 4.197e-05 | 0.001135 |
| 173 | POSITIVE REGULATION OF CELLULAR COMPONENT ORGANIZATION | 9 | 1152 | 4.302e-05 | 0.001157 |
| 174 | PROTEIN ACETYLATION | 4 | 129 | 4.541e-05 | 0.001214 |
| 175 | VENTRICULAR CARDIAC MUSCLE TISSUE DEVELOPMENT | 3 | 45 | 4.578e-05 | 0.001217 |
| 176 | NEGATIVE REGULATION OF CELL PROLIFERATION | 7 | 643 | 4.609e-05 | 0.001219 |
| 177 | REGULATION OF CELL DEATH | 10 | 1472 | 4.783e-05 | 0.001257 |
| 178 | HISTONE H3 ACETYLATION | 3 | 46 | 4.892e-05 | 0.001279 |
| 179 | INTERSPECIES INTERACTION BETWEEN ORGANISMS | 7 | 662 | 5.543e-05 | 0.001433 |
| 180 | SYMBIOSIS ENCOMPASSING MUTUALISM THROUGH PARASITISM | 7 | 662 | 5.543e-05 | 0.001433 |
| 181 | GLOMERULUS DEVELOPMENT | 3 | 49 | 5.919e-05 | 0.001513 |
| 182 | CARDIAC CELL DEVELOPMENT | 3 | 49 | 5.919e-05 | 0.001513 |
| 183 | MUSCLE ORGAN DEVELOPMENT | 5 | 277 | 6.218e-05 | 0.001581 |
| 184 | CELLULAR RESPONSE TO OXYGEN LEVELS | 4 | 143 | 6.786e-05 | 0.001716 |
| 185 | INTERACTION WITH SYMBIONT | 3 | 52 | 7.078e-05 | 0.001771 |
| 186 | POSITIVE REGULATION OF STRIATED MUSCLE CELL DIFFERENTIATION | 3 | 52 | 7.078e-05 | 0.001771 |
| 187 | NEURAL TUBE DEVELOPMENT | 4 | 149 | 7.96e-05 | 0.00197 |
| 188 | CARDIAC MUSCLE TISSUE MORPHOGENESIS | 3 | 54 | 7.927e-05 | 0.00197 |
| 189 | POSITIVE REGULATION OF CHROMOSOME ORGANIZATION | 4 | 150 | 8.169e-05 | 0.002001 |
| 190 | CHROMATIN REMODELING | 4 | 150 | 8.169e-05 | 0.002001 |
| 191 | NEGATIVE REGULATION OF MUSCLE CELL DIFFERENTIATION | 3 | 55 | 8.376e-05 | 0.00203 |
| 192 | REGULATION OF MYOTUBE DIFFERENTIATION | 3 | 55 | 8.376e-05 | 0.00203 |
| 193 | REGULATION OF CHROMATIN ORGANIZATION | 4 | 152 | 8.6e-05 | 0.002073 |
| 194 | POSITIVE REGULATION OF MUSCLE TISSUE DEVELOPMENT | 3 | 56 | 8.841e-05 | 0.00211 |
| 195 | OUTFLOW TRACT MORPHOGENESIS | 3 | 56 | 8.841e-05 | 0.00211 |
| 196 | POSITIVE REGULATION OF DEVELOPMENTAL GROWTH | 4 | 156 | 9.51e-05 | 0.002246 |
| 197 | POSITIVE REGULATION OF CELL PROJECTION ORGANIZATION | 5 | 303 | 9.496e-05 | 0.002246 |
| 198 | PROTEIN ACYLATION | 4 | 159 | 0.0001024 | 0.002406 |
| 199 | POSITIVE REGULATION OF CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 4 | 162 | 0.00011 | 0.002573 |
| 200 | CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 6 | 513 | 0.000118 | 0.002745 |
| 201 | EMBRYONIC HEART TUBE MORPHOGENESIS | 3 | 62 | 0.0001199 | 0.002775 |
| 202 | REGULATION OF PROTEIN BINDING | 4 | 168 | 0.0001266 | 0.002836 |
| 203 | LOOP OF HENLE DEVELOPMENT | 2 | 11 | 0.0001268 | 0.002836 |
| 204 | ENDOCARDIUM DEVELOPMENT | 2 | 11 | 0.0001268 | 0.002836 |
| 205 | NEGATIVE REGULATION OF SPROUTING ANGIOGENESIS | 2 | 11 | 0.0001268 | 0.002836 |
| 206 | POSITIVE REGULATION OF RECEPTOR BIOSYNTHETIC PROCESS | 2 | 11 | 0.0001268 | 0.002836 |
| 207 | EPITHELIAL TO MESENCHYMAL TRANSITION INVOLVED IN ENDOCARDIAL CUSHION FORMATION | 2 | 11 | 0.0001268 | 0.002836 |
| 208 | ENDOTHELIAL TUBE MORPHOGENESIS | 2 | 11 | 0.0001268 | 0.002836 |
| 209 | EYE DEVELOPMENT | 5 | 326 | 0.0001338 | 0.00298 |
| 210 | CELLULAR RESPONSE TO RETINOIC ACID | 3 | 65 | 0.000138 | 0.003058 |
| 211 | CARDIAC LEFT VENTRICLE MORPHOGENESIS | 2 | 12 | 0.000152 | 0.003274 |
| 212 | POSITIVE REGULATION BY HOST OF VIRAL TRANSCRIPTION | 2 | 12 | 0.000152 | 0.003274 |
| 213 | CARDIAC CONDUCTION SYSTEM DEVELOPMENT | 2 | 12 | 0.000152 | 0.003274 |
| 214 | AMYLOID PRECURSOR PROTEIN METABOLIC PROCESS | 2 | 12 | 0.000152 | 0.003274 |
| 215 | DISTAL TUBULE DEVELOPMENT | 2 | 12 | 0.000152 | 0.003274 |
| 216 | LATERAL VENTRICLE DEVELOPMENT | 2 | 12 | 0.000152 | 0.003274 |
| 217 | CHROMATIN MODIFICATION | 6 | 539 | 0.0001545 | 0.003314 |
| 218 | REGULATION OF CELL MORPHOGENESIS INVOLVED IN DIFFERENTIATION | 5 | 337 | 0.0001563 | 0.003336 |
| 219 | MUSCLE ORGAN MORPHOGENESIS | 3 | 70 | 0.000172 | 0.003655 |
| 220 | NEURONAL STEM CELL DIVISION | 2 | 13 | 0.0001794 | 0.003694 |
| 221 | EYELID DEVELOPMENT IN CAMERA TYPE EYE | 2 | 13 | 0.0001794 | 0.003694 |
| 222 | LEFT RIGHT AXIS SPECIFICATION | 2 | 13 | 0.0001794 | 0.003694 |
| 223 | CARDIOBLAST DIFFERENTIATION | 2 | 13 | 0.0001794 | 0.003694 |
| 224 | NEGATIVE REGULATION OF EPIDERMAL CELL DIFFERENTIATION | 2 | 13 | 0.0001794 | 0.003694 |
| 225 | NEUROBLAST DIVISION | 2 | 13 | 0.0001794 | 0.003694 |
| 226 | POSITIVE REGULATION OF GLUCONEOGENESIS | 2 | 13 | 0.0001794 | 0.003694 |
| 227 | EPITHELIAL CELL DEVELOPMENT | 4 | 186 | 0.0001872 | 0.003821 |
| 228 | REGULATION OF CELL PROJECTION ORGANIZATION | 6 | 558 | 0.0001865 | 0.003821 |
| 229 | POSITIVE REGULATION OF PROTEIN BINDING | 3 | 73 | 0.0001949 | 0.003942 |
| 230 | EMBRYONIC HEART TUBE DEVELOPMENT | 3 | 73 | 0.0001949 | 0.003942 |
| 231 | REGULATION OF CYTOKINE PRODUCTION | 6 | 563 | 0.0001958 | 0.003944 |
| 232 | BETA AMYLOID METABOLIC PROCESS | 2 | 14 | 0.0002091 | 0.004195 |
| 233 | ODONTOGENESIS OF DENTIN CONTAINING TOOTH | 3 | 75 | 0.0002111 | 0.004216 |
| 234 | POSITIVE REGULATION OF GENE EXPRESSION EPIGENETIC | 3 | 78 | 0.0002371 | 0.004635 |
| 235 | RESPONSE TO MURAMYL DIPEPTIDE | 2 | 15 | 0.0002411 | 0.004635 |
| 236 | ENDOCARDIAL CUSHION FORMATION | 2 | 15 | 0.0002411 | 0.004635 |
| 237 | RENAL TUBULE DEVELOPMENT | 3 | 78 | 0.0002371 | 0.004635 |
| 238 | ECTODERMAL PLACODE DEVELOPMENT | 2 | 15 | 0.0002411 | 0.004635 |
| 239 | REGULATION OF CELL PROLIFERATION INVOLVED IN HEART MORPHOGENESIS | 2 | 15 | 0.0002411 | 0.004635 |
| 240 | RESPIRATORY SYSTEM DEVELOPMENT | 4 | 197 | 0.0002333 | 0.004635 |
| 241 | ECTODERMAL PLACODE MORPHOGENESIS | 2 | 15 | 0.0002411 | 0.004635 |
| 242 | ECTODERMAL PLACODE FORMATION | 2 | 15 | 0.0002411 | 0.004635 |
| 243 | POSITIVE REGULATION OF MOLECULAR FUNCTION | 10 | 1791 | 0.0002474 | 0.004736 |
| 244 | POSITIVE REGULATION OF PROTEIN MODIFICATION PROCESS | 8 | 1135 | 0.0002564 | 0.004889 |
| 245 | REGULATION OF SYMBIOSIS ENCOMPASSING MUTUALISM THROUGH PARASITISM | 4 | 205 | 0.0002715 | 0.005156 |
| 246 | CARDIAC RIGHT VENTRICLE MORPHOGENESIS | 2 | 16 | 0.0002753 | 0.005165 |
| 247 | NEGATIVE REGULATION OF EPIDERMIS DEVELOPMENT | 2 | 16 | 0.0002753 | 0.005165 |
| 248 | APOPTOTIC PROCESS INVOLVED IN MORPHOGENESIS | 2 | 16 | 0.0002753 | 0.005165 |
| 249 | REGULATION OF TRANSMEMBRANE RECEPTOR PROTEIN SERINE THREONINE KINASE SIGNALING PATHWAY | 4 | 207 | 0.0002817 | 0.005264 |
| 250 | REGULATION OF ERBB SIGNALING PATHWAY | 3 | 83 | 0.0002849 | 0.005302 |
| 251 | POSITIVE REGULATION OF MUSCLE CELL DIFFERENTIATION | 3 | 84 | 0.0002951 | 0.00547 |
| 252 | CELL CELL ADHESION | 6 | 608 | 0.0002964 | 0.005472 |
| 253 | REGULATION OF CHROMATIN BINDING | 2 | 17 | 0.0003117 | 0.005709 |
| 254 | MEMBRANE PROTEIN INTRACELLULAR DOMAIN PROTEOLYSIS | 2 | 17 | 0.0003117 | 0.005709 |
| 255 | GLAND DEVELOPMENT | 5 | 395 | 0.000326 | 0.005949 |
| 256 | NEGATIVE REGULATION OF MYOTUBE DIFFERENTIATION | 2 | 18 | 0.0003503 | 0.006342 |
| 257 | PERICARDIUM DEVELOPMENT | 2 | 18 | 0.0003503 | 0.006342 |
| 258 | NEGATIVE REGULATION OF CELL COMMUNICATION | 8 | 1192 | 0.0003576 | 0.00645 |
| 259 | EMBRYONIC ORGAN DEVELOPMENT | 5 | 406 | 0.0003698 | 0.006644 |
| 260 | REGULATION OF NEURON PROJECTION DEVELOPMENT | 5 | 408 | 0.0003782 | 0.006769 |
| 261 | KIDNEY VASCULATURE DEVELOPMENT | 2 | 19 | 0.0003911 | 0.006867 |
| 262 | REGULATION OF CELL MIGRATION INVOLVED IN SPROUTING ANGIOGENESIS | 2 | 19 | 0.0003911 | 0.006867 |
| 263 | POSITIVE REGULATION OF CARDIAC MUSCLE CELL PROLIFERATION | 2 | 19 | 0.0003911 | 0.006867 |
| 264 | RENAL SYSTEM VASCULATURE DEVELOPMENT | 2 | 19 | 0.0003911 | 0.006867 |
| 265 | POSITIVE REGULATION OF VIRAL PROCESS | 3 | 92 | 0.0003857 | 0.006867 |
| 266 | NEPHRON EPITHELIUM DEVELOPMENT | 3 | 93 | 0.0003981 | 0.006938 |
| 267 | REGULATION OF DNA BINDING | 3 | 93 | 0.0003981 | 0.006938 |
| 268 | TELENCEPHALON DEVELOPMENT | 4 | 228 | 0.0004064 | 0.007057 |
| 269 | REGULATION OF GENE EXPRESSION EPIGENETIC | 4 | 229 | 0.0004132 | 0.007148 |
| 270 | TONGUE DEVELOPMENT | 2 | 20 | 0.0004342 | 0.007427 |
| 271 | MYELOID DENDRITIC CELL DIFFERENTIATION | 2 | 20 | 0.0004342 | 0.007427 |
| 272 | POSITIVE REGULATION OF NEURON PROJECTION DEVELOPMENT | 4 | 232 | 0.000434 | 0.007427 |
| 273 | MEMORY | 3 | 98 | 0.0004642 | 0.007883 |
| 274 | REGULATION OF CANONICAL WNT SIGNALING PATHWAY | 4 | 236 | 0.000463 | 0.007883 |
| 275 | CHROMATIN ORGANIZATION | 6 | 663 | 0.0004704 | 0.007959 |
| 276 | LEFT RIGHT PATTERN FORMATION | 2 | 21 | 0.0004794 | 0.008024 |
| 277 | REGULATION OF RECEPTOR BIOSYNTHETIC PROCESS | 2 | 21 | 0.0004794 | 0.008024 |
| 278 | APOPTOTIC PROCESS INVOLVED IN DEVELOPMENT | 2 | 21 | 0.0004794 | 0.008024 |
| 279 | MODIFICATION OF MORPHOLOGY OR PHYSIOLOGY OF OTHER ORGANISM | 3 | 100 | 0.0004925 | 0.008213 |
| 280 | KERATINOCYTE DIFFERENTIATION | 3 | 101 | 0.000507 | 0.008395 |
| 281 | CELL PROLIFERATION | 6 | 672 | 0.0005052 | 0.008395 |
| 282 | REGULATION OF ORGAN MORPHOGENESIS | 4 | 242 | 0.0005089 | 0.008397 |
| 283 | ENDOCARDIAL CUSHION MORPHOGENESIS | 2 | 22 | 0.0005268 | 0.008601 |
| 284 | REGULATION OF ANDROGEN RECEPTOR SIGNALING PATHWAY | 2 | 22 | 0.0005268 | 0.008601 |
| 285 | HISTONE H3 DEACETYLATION | 2 | 22 | 0.0005268 | 0.008601 |
| 286 | REGULATION OF MUSCLE TISSUE DEVELOPMENT | 3 | 103 | 0.0005369 | 0.008705 |
| 287 | REGULATION OF MUSCLE ORGAN DEVELOPMENT | 3 | 103 | 0.0005369 | 0.008705 |
| 288 | ODONTOGENESIS | 3 | 105 | 0.0005679 | 0.009176 |
| 289 | REGULATION OF EPIDERMAL GROWTH FACTOR ACTIVATED RECEPTOR ACTIVITY | 2 | 23 | 0.0005764 | 0.009185 |
| 290 | POSITIVE REGULATION OF MULTICELLULAR ORGANISMAL METABOLIC PROCESS | 2 | 23 | 0.0005764 | 0.009185 |
| 291 | POSITIVE REGULATION OF TRANSCRIPTION FROM RNA POLYMERASE II PROMOTER IN RESPONSE TO STRESS | 2 | 23 | 0.0005764 | 0.009185 |
| 292 | POSITIVE REGULATION OF COLLAGEN METABOLIC PROCESS | 2 | 23 | 0.0005764 | 0.009185 |
| 293 | SPINAL CORD DEVELOPMENT | 3 | 106 | 0.0005839 | 0.009272 |
| 294 | RESPONSE TO RETINOIC ACID | 3 | 107 | 0.0006001 | 0.009497 |
| 295 | NEGATIVE REGULATION OF BLOOD VESSEL ENDOTHELIAL CELL MIGRATION | 2 | 24 | 0.0006282 | 0.009744 |
| 296 | POSITIVE REGULATION OF CELLULAR RESPONSE TO TRANSFORMING GROWTH FACTOR BETA STIMULUS | 2 | 24 | 0.0006282 | 0.009744 |
| 297 | POSITIVE REGULATION OF TRANSFORMING GROWTH FACTOR BETA RECEPTOR SIGNALING PATHWAY | 2 | 24 | 0.0006282 | 0.009744 |
| 298 | NEGATIVE REGULATION OF MYOBLAST DIFFERENTIATION | 2 | 24 | 0.0006282 | 0.009744 |
| 299 | EPITHELIAL CELL DIFFERENTIATION INVOLVED IN KIDNEY DEVELOPMENT | 2 | 24 | 0.0006282 | 0.009744 |
| 300 | CARDIAC EPITHELIAL TO MESENCHYMAL TRANSITION | 2 | 24 | 0.0006282 | 0.009744 |
| 301 | SINGLE ORGANISM CELL ADHESION | 5 | 459 | 0.0006465 | 0.009994 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | NOTCH BINDING | 8 | 18 | 5.389e-19 | 5.006e-16 |
| 2 | TRANSCRIPTION FACTOR ACTIVITY PROTEIN BINDING | 12 | 588 | 3.162e-11 | 1.469e-08 |
| 3 | TRANSCRIPTION COACTIVATOR ACTIVITY | 9 | 296 | 4.573e-10 | 1.416e-07 |
| 4 | TRANSCRIPTION FACTOR BINDING | 10 | 524 | 3.784e-09 | 8.789e-07 |
| 5 | CHROMATIN BINDING | 9 | 435 | 1.323e-08 | 2.458e-06 |
| 6 | RNA POLYMERASE II TRANSCRIPTION FACTOR BINDING | 5 | 104 | 5.265e-07 | 8.153e-05 |
| 7 | RECEPTOR BINDING | 12 | 1476 | 9.393e-07 | 0.0001247 |
| 8 | CORE PROMOTER BINDING | 5 | 152 | 3.439e-06 | 0.0003549 |
| 9 | PEPTIDE N ACETYLTRANSFERASE ACTIVITY | 4 | 67 | 3.381e-06 | 0.0003549 |
| 10 | BETA CATENIN BINDING | 4 | 84 | 8.355e-06 | 0.0006381 |
| 11 | CALCIUM ION BINDING | 8 | 697 | 8.095e-06 | 0.0006381 |
| 12 | RNA POLYMERASE II REPRESSING TRANSCRIPTION FACTOR BINDING | 3 | 27 | 9.617e-06 | 0.0006381 |
| 13 | CHROMATIN DNA BINDING | 4 | 80 | 6.878e-06 | 0.0006381 |
| 14 | N ACETYLTRANSFERASE ACTIVITY | 4 | 87 | 9.607e-06 | 0.0006381 |
| 15 | NF KAPPAB BINDING | 3 | 30 | 1.331e-05 | 0.0008241 |
| 16 | ACETYLTRANSFERASE ACTIVITY | 4 | 103 | 1.876e-05 | 0.001065 |
| 17 | N ACYLTRANSFERASE ACTIVITY | 4 | 104 | 1.949e-05 | 0.001065 |
| 18 | MACROMOLECULAR COMPLEX BINDING | 10 | 1399 | 3.091e-05 | 0.001566 |
| 19 | STRUCTURE SPECIFIC DNA BINDING | 4 | 118 | 3.203e-05 | 0.001566 |
| 20 | TRANSCRIPTION FACTOR ACTIVITY RNA POLYMERASE II TRANSCRIPTION FACTOR BINDING | 4 | 133 | 5.116e-05 | 0.002376 |
| 21 | ACTIVATING TRANSCRIPTION FACTOR BINDING | 3 | 57 | 9.322e-05 | 0.003936 |
| 22 | REPRESSING TRANSCRIPTION FACTOR BINDING | 3 | 57 | 9.322e-05 | 0.003936 |
| 23 | HISTONE DEACETYLASE ACTIVITY H3 K14 SPECIFIC | 2 | 12 | 0.000152 | 0.006139 |
| 24 | SMAD BINDING | 3 | 72 | 0.000187 | 0.007145 |
| 25 | ENZYME BINDING | 10 | 1737 | 0.0001923 | 0.007145 |
| 26 | REGULATORY REGION NUCLEIC ACID BINDING | 7 | 818 | 0.0002075 | 0.007415 |
| 27 | PEROXISOME PROLIFERATOR ACTIVATED RECEPTOR BINDING | 2 | 15 | 0.0002411 | 0.008102 |
| 28 | CORE PROMOTER PROXIMAL REGION DNA BINDING | 5 | 371 | 0.0002442 | 0.008102 |
| 29 | TRANSFERASE ACTIVITY TRANSFERRING ACYL GROUPS OTHER THAN AMINO ACYL GROUPS | 4 | 211 | 0.000303 | 0.009478 |
| 30 | TRANSCRIPTIONAL REPRESSOR ACTIVITY RNA POLYMERASE II TRANSCRIPTION FACTOR BINDING | 3 | 86 | 0.0003163 | 0.009478 |
| 31 | NAD DEPENDENT PROTEIN DEACETYLASE ACTIVITY | 2 | 17 | 0.0003117 | 0.009478 |
| Num | GO | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|
| 1 | CHROMOSOME | 10 | 880 | 4.933e-07 | 0.000144 |
| 2 | CHROMATIN | 8 | 441 | 2.651e-07 | 0.000144 |
| 3 | NUCLEOPLASM PART | 9 | 708 | 8.31e-07 | 0.0001618 |
| 4 | NUCLEAR CHROMATIN | 6 | 291 | 4.885e-06 | 0.0007133 |
| 5 | GOLGI MEMBRANE | 8 | 703 | 8.618e-06 | 0.001007 |
| 6 | APICAL PART OF CELL | 6 | 361 | 1.668e-05 | 0.001392 |
| 7 | ACETYLTRANSFERASE COMPLEX | 4 | 100 | 1.669e-05 | 0.001392 |
| 8 | GOLGI APPARATUS PART | 8 | 893 | 4.828e-05 | 0.003524 |
| 9 | TRANSCRIPTION FACTOR COMPLEX | 5 | 298 | 8.781e-05 | 0.005128 |
| 10 | APICAL PLASMA MEMBRANE | 5 | 292 | 7.978e-05 | 0.005128 |
| 11 | NUCLEAR CHROMOSOME | 6 | 523 | 0.0001311 | 0.006961 |
| 12 | SIN3 COMPLEX | 2 | 13 | 0.0001794 | 0.008733 |
| 13 | TRANSCRIPTIONAL REPRESSOR COMPLEX | 3 | 74 | 0.0002029 | 0.009114 |
| 14 | ADA2 GCN5 ADA3 TRANSCRIPTION ACTIVATOR COMPLEX | 2 | 15 | 0.0002411 | 0.009564 |
| 15 | GOLGI APPARATUS | 9 | 1445 | 0.0002456 | 0.009564 |
Over-represented Pathway
| Num | Pathway | Pathview | Overlap | Size | P Value | Adj. P Value |
|---|---|---|---|---|---|---|
| 1 | hsa04330_Notch_signaling_pathway | 31 | 47 | 5.891e-88 | 1.06e-85 | |
| 2 | hsa04320_Dorso.ventral_axis_formation | 4 | 25 | 5.838e-08 | 5.254e-06 | |
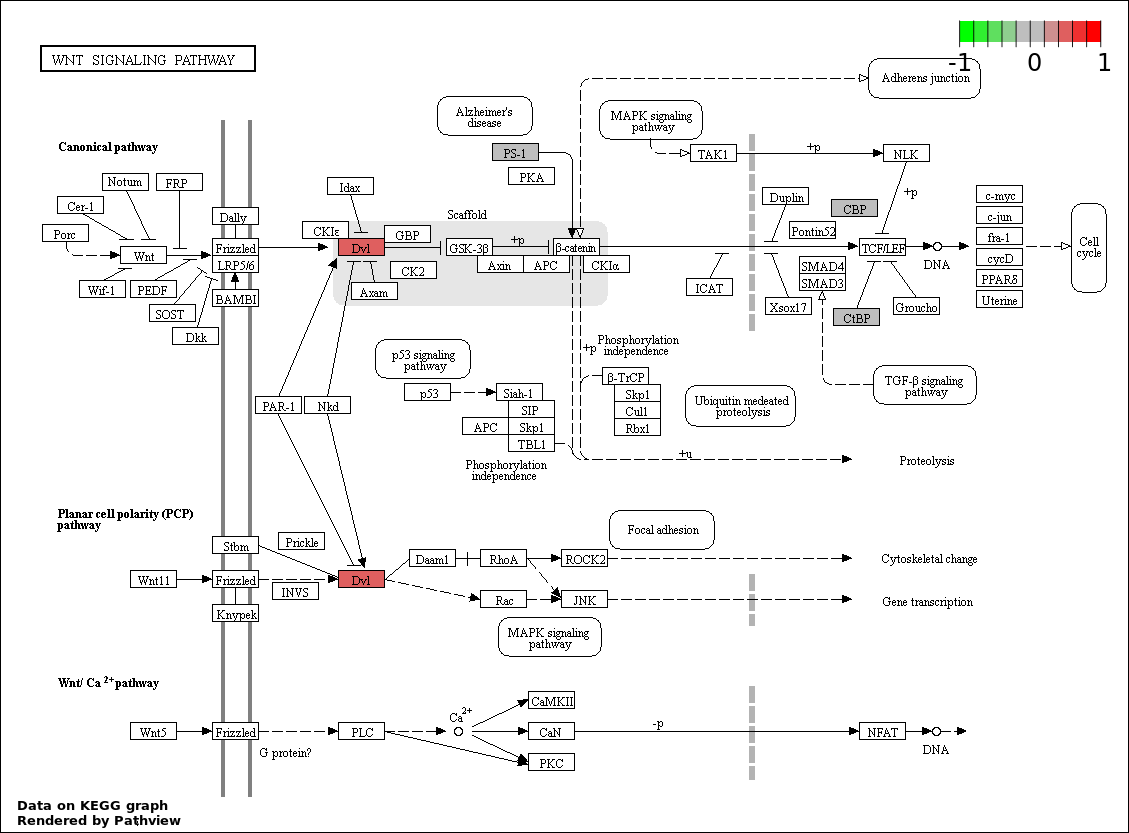
| 3 | hsa04310_Wnt_signaling_pathway | 5 | 151 | 3.329e-06 | 0.0001997 | |
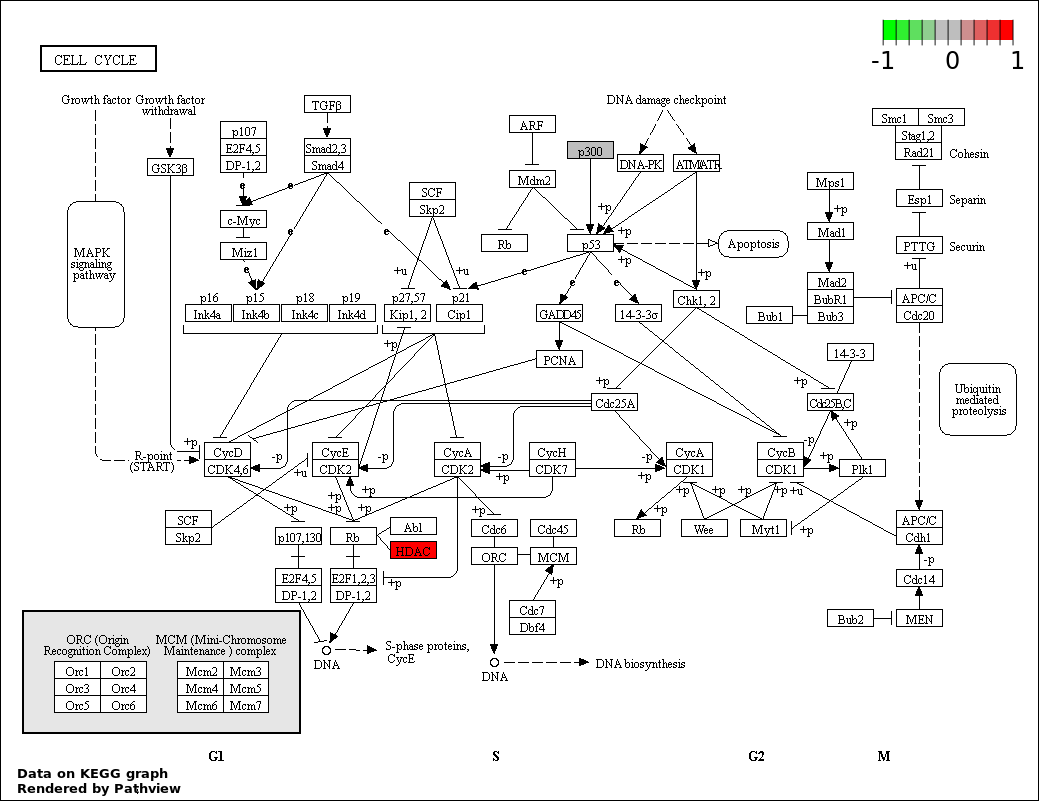
| 4 | hsa04110_Cell_cycle | 4 | 128 | 4.405e-05 | 0.001982 | |
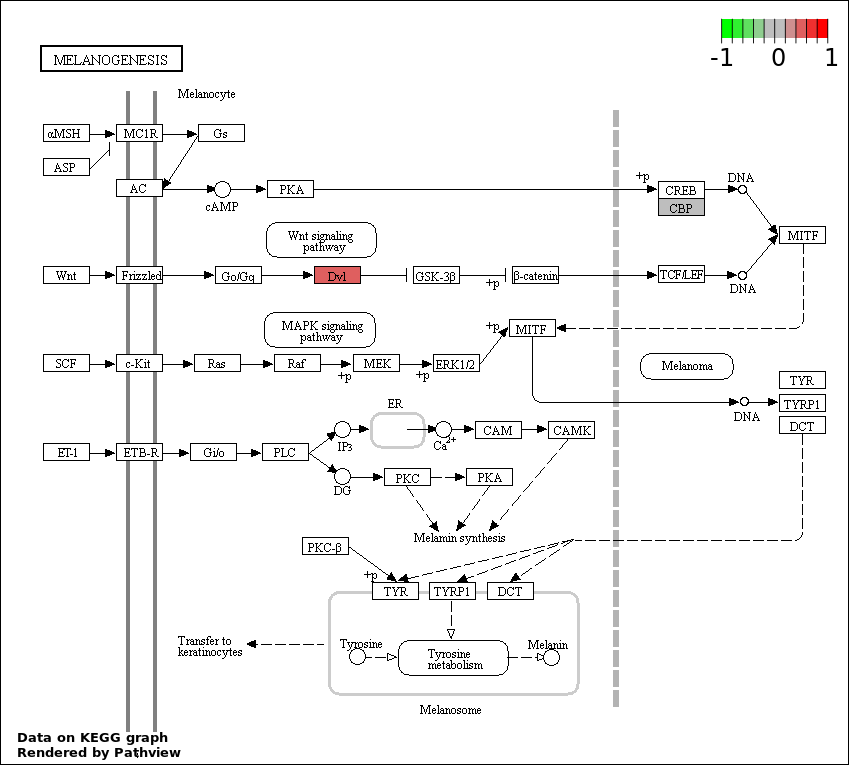
| 5 | hsa04916_Melanogenesis | 3 | 101 | 0.000507 | 0.01825 | |
| 6 | hsa00514_Other_types_of_O.glycan_biosynthesis | 2 | 46 | 0.002306 | 0.06919 | |
| 7 | hsa04720_Long.term_potentiation | 2 | 70 | 0.005259 | 0.1284 | |
| 8 | hsa04520_Adherens_junction | 2 | 73 | 0.005706 | 0.1284 | |
| 9 | hsa04350_TGF.beta_signaling_pathway | 2 | 85 | 0.007662 | 0.1532 | |
| 10 | hsa04722_Neurotrophin_signaling_pathway | 2 | 127 | 0.0165 | 0.2969 | |
| 11 | hsa04630_Jak.STAT_signaling_pathway | 2 | 155 | 0.02396 | 0.392 |





lncRNA-mediated sponge
| Num | lncRNA | miRNAs | miRNAs count | Gene | Sponge regulatory network | lncRNA log2FC | lncRNA pvalue | Gene log2FC | Gene pvalue | lncRNA-gene Pearson correlation |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | AGAP11 |
hsa-miR-106b-5p;hsa-miR-17-5p;hsa-miR-181a-5p;hsa-miR-181b-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-25-3p;hsa-miR-590-5p;hsa-miR-93-5p | 11 | KAT2B | Sponge network | -1.728 | 0.00016 | -1.911 | 0 | 0.441 |
| 2 | AGAP11 |
hsa-miR-103a-3p;hsa-miR-107;hsa-miR-130b-5p;hsa-miR-15a-5p;hsa-miR-15b-5p;hsa-miR-16-1-3p;hsa-miR-16-5p;hsa-miR-17-5p;hsa-miR-18a-3p;hsa-miR-200a-5p;hsa-miR-200b-5p;hsa-miR-25-3p;hsa-miR-423-5p;hsa-miR-424-5p;hsa-miR-590-5p | 15 | NOTCH2 | Sponge network | -1.728 | 0.00016 | -0.258 | 0.28775 | 0.35 |
| 3 | PCA3 |
hsa-miR-106b-5p;hsa-miR-17-5p;hsa-miR-181a-5p;hsa-miR-181b-5p;hsa-miR-19a-3p;hsa-miR-19b-3p;hsa-miR-20a-3p;hsa-miR-20a-5p;hsa-miR-25-3p;hsa-miR-590-3p;hsa-miR-590-5p;hsa-miR-93-5p | 12 | KAT2B | Sponge network | -2.778 | 8.0E-5 | -1.911 | 0 | 0.339 |